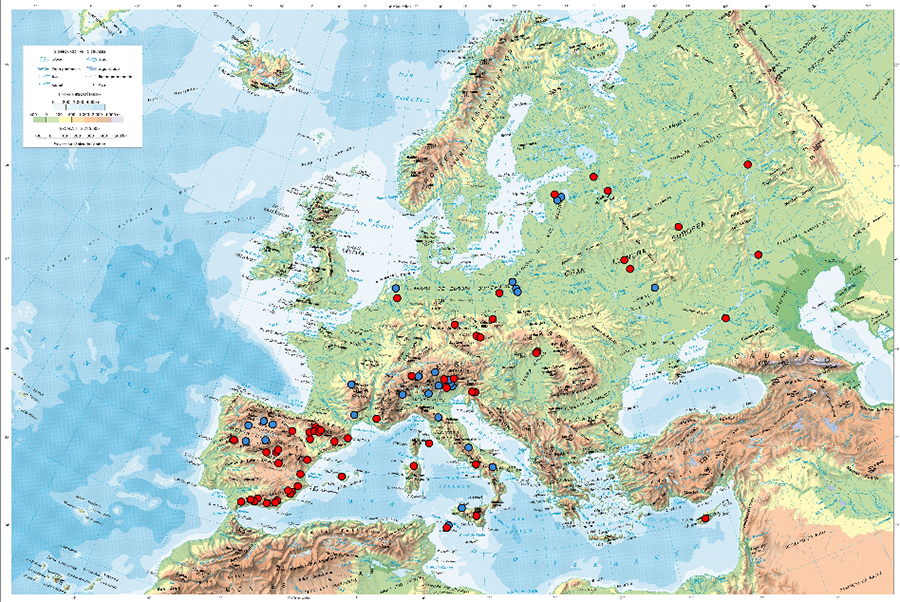
En relación con el CBA, el hecho de que este mostrara
valores elevados en bosques desarrollados en zonas de clima continental húmedo
de verano cálido (Dfb) responde a la relación positiva y significativa
existente entre las tasas de crecimiento de los árboles durante la sucesión
secundaria, la humedad y la temperatura, registrada a escala global (Anderson et al., 2006). El CBA registró su máximo en
los bosques de coníferas desarrollados en pastizales abandonados de los Alpes
italianos (400 Mg C ∙ ha-1; Thuille y
Schulze, 2006), 75-100 años después del abandono. También alcanzó valores
elevados (250-270 Mg C ∙ ha-1), tras un periodo similar, en bosques de coníferas (Bose et al., 2014), en bosques mixtos de coníferas y
especies de hoja caduca (Weissgeber et al., 2024),
y en bosques de especies de hoja caduca (Alberti et al.,
2008), desarrollados en tierras agrarias abandonadas de los Países Bajos,
el Macizo Central francés y los Apeninos, respectivamente.

Figura 7.
Aspecto de los nuevos bosques desarrollados en tierras agrarias abandonadas en Europa: bosque dominado por Betula pendula Roth.
desarrollado en antiguos campos de cultivo (Región de Moscú, Rusia (a))
bosque dominado por Quercus pyrenaica Willd. desarrollado en un antiguo
cultivo de fresas en Linares de Riofrío (Salamanca, España), 41-62 años después
del abandono (b) (la línea discontinua indica el trazo del antiguo muro
de piedra seca, aún distinguible, que delimitaba la parcela), bosque dominado
por Ostrya carpinifolia Scopoli, 1772 y Q. humilis Mill.
desarrollado en una antigua pradera de siega en el fondo de una dolina kárstica
(Podgorski Kras, Eslovenia), 25 años después del abandono (c). Fotos: Irina Kurganova, el autor, Mateja B. Valjavec.
Figure 7. Appearance
of new forests developed on abandoned agricultural land in Europe: a forest
dominated by Betula pendula Roth. developing on former farmland (Moscow Region,
Russia (a)), a forest dominated by Quercus pyrenaica
Willd. developed on a former strawberry field in Linares de Riofrío (Salamanca,
Spain), 41–62 years after abandonment (b) (the
dotted line indicates the outline of the former dry stone wall, still
distinguishable, which marked the boundary of the plot), forest dominated by
Ostrya carpinifolia Scopoli, 1772 and Q. humilis Mill. developed on a former
hay meadow at the bottom of a karst doline (Podgorski Kras, Slovenia), 25 years
after abandonment (c). Photos: Irina
Kurganova, the author, Mateja B. Valjavec.
El escaso número de trabajos que tuvieron en cuenta el CBS
contrasta con la importancia de dicho compartimento para la acumulación de C
durante la sucesión secundaria (Clemmensen et al.,
2013) y está relacionado con las dificultades para realizar mediciones
directas de dicha variable. De hecho, en los trabajos que analizaron el CBS,
este fue estimado mediante el uso de ecuaciones alométricas que permitían el
cálculo de la BS a partir de medidas dendrométricas de la parte aérea de los
árboles (ej. Ruiz-Peinado et al., 2012). El CBS
alcanzó valores entre 21 y 62 Mg C ∙ ha-1 (Alberti
et al., 2008; Uri et al., 2012; Velázquez et al., 2023).
En ambiente Mediterráneo, y entre 40 y 65 años después del abandono, la
proporción de C acumulado en la BS respecto a la BA era más alta en zonas con
PMA < 1000 mm (35 %; Velázquez et al., 2023)
que en zonas con PMA > 1000 mm (26 %; Pellis et al.,
2019), lo que se relaciona con la mayor alocación de recursos en las raíces
en las zonas de menor precipitación para hacer frente a los periodos de sequía
estival.
El escaso número de trabajos que tuvieron en cuenta la CMM
puede estar relacionado con el hecho de que esta se genera debido a la
mortalidad de los primeros árboles que colonizan las tierras agrarias
abandonadas, lo que suele tener lugar tras un periodo de tiempo mayor (~100
años; Hooker y Compton, 2003) que el periodo
medio considerado en los estudios analizados (82 años). Sea como fuere, las reservas de C en la MM y la HOJ alcanzaron
valores máximos de 12-14 Mg C ∙ ha-1 entre 60 y 100 años después del
abandono (Alberti et al., 2008; Badalamenti et al., 2019; Pellis
et al., 2019), lo que les sitúa como compartimentos menores desde el punto
de vista de la acumulación de C.
Patrones de cambio en la
acumulación de C
Los patrones de cambio registrados en el COS fueron
enormemente variables. La mayor parte de los trabajos que registraron un
aumento en el COS se desarrollaron en zonas que partían de un bajo nivel
inicial (ej. Badalamenti et al., 2019; Kurganova et al., 2019;
Company et al., 2022; Cortijos-López
et al., 2023). En los suelos de bosques caducifolios desarrollados en
tierras agrarias abandonadas en los Apeninos (Italia; Chiti
et al., 2018) y la Región de Moscú (Rusia; Kurganova et al., 2019), 64 y 35 años después
del abandono, el COS fue un 120 y un 134 % mayor, respectivamente, que el
registrado en campos contiguos aún en uso. En los bosques dominados por Quercus
ilex de la Isla Pantelleria (Italia), > 100 años después del abandono,
el COS era un 200 % mayor que en las zonas de matorral (maquia)
desarrollados en etapas iniciales (Badalamenti et
al., 2019). El aumento del COS durante la colonización vegetal de tierras
agrarias abandonadas viene explicado por la mayor protección del suelo frente a
la erosión, la reducción de la descomposición de la MO al desaparecer el
laboreo (Marzaioli et al., 2010; Rhyzova et al., 2020) y el aumento de los aportes de MO
procedentes de residuos vegetales de especies herbáceas (Vaccari
et al., 2012; Bohner et al., 2019).
Algunos autores también señalaron la existencia de un
aumento del COS en las etapas iniciales de la sucesión secundaria en tierras
agrarias abandonadas seguido de una estabilización (Velázquez
et al., 2023) o incluso una disminución (ej. Pellis
et al., 2019; Ovsepyan et al., 2019; Bell et al., 2021) en las etapas finales. Por ejemplo, en
los Phaeozems de los bosques deciduos desarrollados en cultivos
abandonados de la Región de Moscú (Rusia), las parcelas abandonadas hace 7 años
registraron una tasa de acumulación del COS de 1.31 Mg C ∙ ha-1 ∙
año-1 y las abandonadas hace 35, una tasa de
0.42 Mg C ∙ ha-1 ∙ año-1 (Ovsepyan
et al., 2019). El hecho de que la acumulación de COS disminuya con la edad
de abandono podría indicar un efecto de saturación del sumidero de C en el
suelo (Stewart et al., 2007), o simplemente, ser el
resultado de una contribución proporcionalmente mayor de los aportes de MO al
suelo durante las etapas iniciales de la sucesión respecto a las etapas finales
(Bell et al., 2021; Velázquez
et al., 2023).
La mayor parte de los trabajos que detectaron una
disminución del COS durante la colonización vegetal en tierras agrarias
abandonadas, se desarrollaron en sistemas como los pastizales de montaña (Alberti et al., 2008, 2011; Nadal-Romero et al., 2016; Oddi
et al., 2025) y las praderas de siega (Rhyzova et
al., 2020), en los que el contenido en C era elevado antes del abandono.
Dicha disminución se explica por la reducción de los aportes MO procedentes de
residuos vegetales de las especies herbáceas y la rápida descomposición de la
MO del suelo durante las primeras etapas de la sucesión en este tipo de
sistemas (Alberti et al., 2008; Rhyzova et al., 2020). La
disminución del COS que se producía en el suelo superficial de los bosques
desarrollados en cultivos abandonados de Polonia Central (Jonczak et al., 2023), venía
explicada, sin embargo, porque la colonización de los abedules (Betula
pendula Roth) provocaba un aumento en la proporción de fracciones de C más
lábiles ó susceptibles a la degradación en la hojarasca, aumentando por tanto
su tasa de mineralización. Algunos de los trabajos que registraron una
disminución del COS tras el abandono encontraron una recuperación posterior a
partir del momento en el que los aportes de residuos vegetales de la vegetación
colonizadora compensaban las pérdidas iniciales (Navas et
al., 2012; Rhyzova et al., 2020). En cultivos de
cereal abandonados de la península ibérica, la disminución de las reservas del
C durante las etapas iniciales de la sucesión puede venir explicada por el
hecho de que, en muchas ocasiones, en los suelos de este tipo de sistemas la
comunidad microbiana es menor y menos diversa que en los bosques adyacentes
debido al efecto negativo de los laboreos frecuentes y el uso de pesticidas (Zornoza et al., 2009), lo que genera a su vez, una
menor tasa de descomposición de la hojarasca y una menor incorporación de C al
suelo (Acuña-Mínguez et al., 2024). Dicha
disminución también puede estar relacionada con el aumento de la erosión tras
el abandono de la actividad agraria (Ruecker et al.,
1999), particularmente en sistemas agrícolas con una baja cobertura
herbácea (ej. almendrales; Rodrigo-Comino et al.
(2018).
Según los trabajos analizados, las reservas de C en el suelo
no alcanzaban valores similares a los existentes en los ecosistemas
semi-naturales cercanos hasta 80 después del abandono en los bosques de
coníferas desarrollados en pastizales alpinos (Thuille
y Schulze, 2006) y hasta 100-150 años después del abandono en la Taiga y la
estepa húmeda (Kalinina et al., 2015). Sin embargo,
en la estepa semiárida de la parte europea de Rusia, la acumulación de C en el
suelo se producía apenas 30 años después del abandono
(Ovsepyan et al., 2019). Se produjera o no dicha recuperación, varios trabajos
indicaron una saturación del sumidero de C en el suelo entre los 28 y 55
años (Alberti et al., 2008; Guidi
et al., 2014; Van Hall et al., 2017; Pellis et al., 2019) o entre 60 y 80 años después del
abandono (Cortijos-López et al., 2023; Velázquez et al., 2023).
En relación con el papel del suelo
profundo y el suelo superficial en la fijación de C, los trabajos analizados
mostraron que, mientras el suelo superficial mostraba mayores tasas de
acumulación, el suelo profundo acumulaba una mayor reserva de C (p. ej. Nóvak et al., 2014; Gabarrón-Galeote et al., 2015;
Nadal-Romero et al., 2016). Dicha reserva, sin embargo, no era necesariamente más
estable, y algunos trabajos mostraron, de hecho, una disminución (Sokolowska et al., 2020),
lo que viene explicado por la reducción de la actividad microbiana en
horizontes profundos.
Aunque la colonización vegetal de
plantas leñosas en tierras agrarias abandonadas tenía un claro efecto de
retraso (lag-effect; Fig. A2 del anexo),
y no se producía hasta unos años después del abandono (p. ej. 15-20; Velázquez et al., 2023). A partir de entonces y
durante las primeras décadas, el CBA seguía un claro patrón de aumento, con una
elevada tasa de acumulación (Uri
et al., 2012; Aosaar
et al., 2016; Velázquez
et al., 2023). Dicha tasa, sin embargo, se
reducía entre 60 y 80 años después del abandono (Bose et
al., 2014, Weissgeber et al., 2024), y 100
años después podía volverse incluso negativa (Thuille
et al., 2000), lo que viene explicado por la existencia de fenómenos de
auto-aclareo.
Aunque la CBS también aumentó (Alberti
et al., 2008; Uri et al., 2012; Velázquez et al., 2023), algunos
autores detectaron una disminución de la importancia relativa de este
compartimento en relación con la BA (p. ej. Bijak
et al., 2013), lo que corrobora la
tendencia general de los árboles a acumular más C en la BA (y menos en la BS)
durante la sucesión secundaria (Law et al., 2003). El
CMM y el CHOJ también aumentaron durante la sucesión secundaria en tierras
agrarias abandonadas (p. ej. Alberti et al., 2008; Pellis et al., 2019; Velázquez
et al., 2023), alcanzando valores máximos 60-100 años después del abandono,
lo que puede explicarse por la existencia de un punto de saturación en la
máxima cantidad de materia orgánica mineralizable (Badalamenti
et al., 2019) o en los aportes orgánicos procedentes del desfronde (Uri et al., 2012).
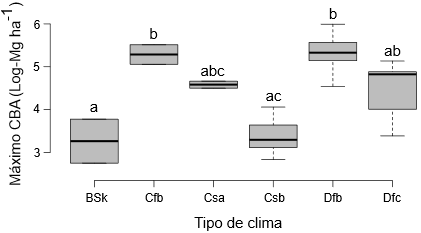
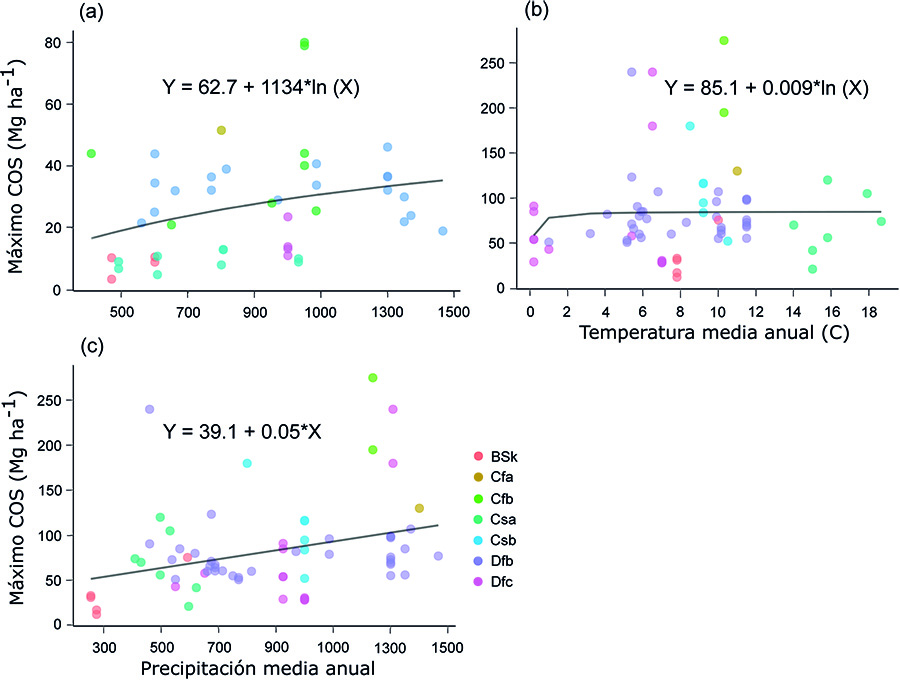
Factores determinantes de la acumulación de C
La relación positiva y significativa entre los valores
máximos del COS en el suelo superficial y la PMA (Fig. 5a)
puede deberse al hecho de que, según aumenta la precipitación y, por tanto, la
humedad del suelo también aumenta la actividad microbiana, produciéndose una
mayor descomposición de la materia orgánica (Zeglin et
al., 2013). Resulta destacable, sin embargo, que los valores máximos del
COS fueran especialmente altos entre 800 y 1100 mm, lo que corrobora los
resultados de trabajos anteriores que indican que la máxima acumulación del COS
se produce en un rango de PMA entre 650 y 900-1000 (Alberti
et al., 2011; Gabarrón-Galeote et al., 2015; Novara et al., 2017) o entre 450 y 900 mm (Bell et al., 2021). El hecho de
que, con valores de PMA > 1300 mm, la pendiente de la curva de
Michaelis-Menten se redujera, sugiere que, en climas excesivamente lluviosos,
se producen pérdidas de C (Thuille y
Schulze, 2006; Pellis
et al., 2019). En contraste, la relación lineal,
positiva y significativa entre los valores máximos de COS en todo el perfil y
la PMA (Fig. 5c)
indica que el COS puede seguir acumulándose en horizontes profundos. La
relación positiva y significativa entre los valores máximos del COS y la TMA en
todo el perfil del suelo (Fig. 5b) por su parte, está en línea con los resultados de Poeplau et al. (2011), Novara et al. (2017) y Pellis et al. (2019). Es
interesante observar el hecho de que, en nuestro caso, los valores máximos del
COS aparecieron con una TMA entre 4 y 12 °C, mientras que en la revisión
efectuada por Bell et al. (2021) para la península ibérica, dichos valores se alcanzaron
con una TMA de 13-17 °C, lo que puede deberse al menor rango de TMA analizado
en este estudio.
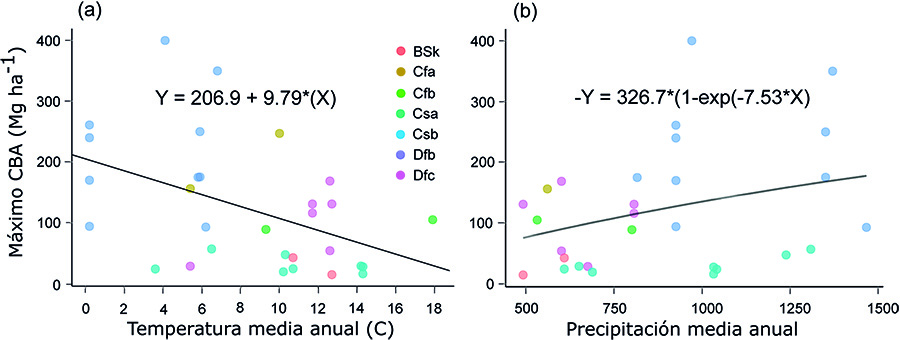
Aunque nuestros resultados indican la existencia de una
relación positiva entre los valores máximos del CBA y la PMA (Fig.
6b; Tabla A3 del
Anexo), es importante destacar que en los nuevos bosques y zonas de
matorral desarrollados en tierras abandonadas en el Centro-Norte de España, el
CBA estaba relacionado de forma negativa con la precipitación media del mes más
seco (Velázquez et al., 2023), lo que puede explicarse por la estrecha relación
entre esta y la CBS, y la importancia de una baja precipitación estival para el
desarrollo de los sistemas radiculares en climas de tipo Mediterráneo
sub-húmedo (Moreno et al., 2011). Estos resultados sugieren que el efecto de la precipitación
en la acumulación de C en el suelo se produce de forma indirecta, a través del
efecto que tiene esta variable en la dinámica de la vegetación (Wang et al., 2020) y los aportes
de materia orgánica (Uri et al., 2012).
En relación con el tipo de uso
agrícola previo al abandono, la capacidad de secuestro de C en el suelo era
mayor en los cultivos leñosos que los cultivos de cereal abandonados (Spohn et al., 2016; Romero-Díaz et al., 2017; Badalamenti et al., 2019)
porque, al encontrarse en zonas de mayor pendiente y suelos someros, apenas
recibían aportes de materia orgánica y por tanto, tenían un nivel inicial de
COS más bajo (Calvo de Anta et al., 2020). Sin embargo, en aquellos sistemas agrarios en los
que los aportes de materia orgánica en forma de abono o residuos vegetales eran
mayores, la acumulación de C a lo largo de la colonización vegetal tras el
abandono era menor y podían producirse incluso pérdidas (ej. Navas et al., 2012). La intensidad de uso también era
importante. En tierras agrarias abandonadas de la República Checa, Kopecky y Vojta (2009) detectaron que el COS
era menor en aquellos suelos en los que el laboreo había sido más intenso. De hecho, las marcas dejadas por el laboreo seguían
distinguiéndose en el perfil de los suelos de los bosques templados
desarrollados en cultivos abandonados de la parte europea de Rusia, más de 100
años después del abandono (Kalinina et al., 2018).
Otro de los legados del uso
agrícola previo es el contenido en N que, según varios de los trabajos
analizados, mostró una relación positiva y significativa con el COS (p. ej. Alberti et al., 2011; Uri et al., 2012; Spohn et al., 2016; Velázquez et al., 2023).
Dicho aumento paralelo entre el COS y el N se producía sobretodo en aquellas
zonas en las que el valor inicial de dicho elemento era menor y, por tanto,
resultaba más limitante para la producción primaria (Rastetter et al., 1992), y también en aquellas en las que se registró una
presencia importante de especies con organismos simbióticos fijadores de N (ej.
Nadal-Romero et al., 2018). En otras zonas, sin embargo, no se detectó una relación
clara entre el contenido de N y el COS (Bell et al.,
2021). En la cronosecuencia desarrollada por Aossar et al. (2016) en campos
abandonados de Estonia, el contenido en N en el suelo superficial aumentó
debido a la presencia de bacterias fijadoras simbióticas en las raíces de los
árboles de la especie dominante (Betula pendula Roth), mientras que el
COS disminuía debido a la menor tasa de descomposición de la hojarasca de dicha
especie. La mayor disponibilidad de N en el suelo de muchas tierras agrarias
abandonadas como legado de las fertilizaciones previas provoca que los árboles
que se desarrollan en estas tengan tasas de crecimiento mayores que las que
presentan en los bosques pre-existentes de zonas adyacentes y, por tanto, una
mayor acumulación de C en la biomasa viva durante la sucesión secundaria (Bose et al., 2014; Vilà-Cabrera et al., 2017;
Alfaro-Sánchez et al., 2019). La influencia del contenido en N en el CBA puede
provocar, a su vez, un efecto positivo indirecto en el COS a través de un
aumento de los aportes de materia orgánica en forma de hojarasca, como ocurre
en los bosques desarrollados en prados de siega abandonados en el fondo de las
dolinas eslovenas (Valjavec et al., 2022; Fig. 7c). De hecho, los aportes de hojarasca pueden explicar el
aumento de la ratio C:N durante la sucesión secundaria en tierras agrarias
abandonadas (Kopecky y Vojta, 2009; Spohn et al., 2016; Bell et al., 2021). Los suelos de muchas tierras agrarias abandonadas
cuentan también con un mayor contenido en P, lo que incrementa la actividad de
enzimas como la fosfatasa y reduce la limitación de este elemento para la
producción primaria (Acuña-Minguez et al.,
2024). Sin embargo, a lo largo de la sucesión
secundaria, el contenido en P en los suelos de tierras agrarias abandonadas
puede disminuir debido a la erosión (p. ej. Ruecker
et al., 1999) o aumentar debido a los aportes de
residuos vegetales con un alto contenido en dicho elemento (Kalinina et al., 2015).
La textura del suelo también
ejercía una influencia clave en la dinámica del COS. En general, este mostraba
un mayor aumento en suelos de textura fina, en los que está más protegido de la
degradación física y química (p. ej. Bose et al.,
2014; Lichner et
al., 2023; Fig. A3
del Anexo). Por el contrario, Velázquez et al. (2023)
registraron un mayor aumento del COS en suelos de textura gruesa. Dichos
suelos, sin embargo, estaban en zonas que habían sido utilizadas como
pastizales durante ciertas épocas del año antes de su abandono, por lo que
habrían contado con un mayor aporte de residuos vegetales.
Muchos trabajos también
mencionaron la densidad de individuos arbóreos como factor determinante para la
acumulación del CBA. En los bosques más maduros, desarrollados en tierras
agrarias abandonadas, los procesos de autoaclareo provocaban una reducción de
la competencia por la luz y los nutrientes y, por tanto, un mayor crecimiento
de los árboles persistentes, lo que daba lugar a una relación negativa entre la
densidad de individuos y la CBA (Uri et al., 2012; Bose et al., 2014; Aossar et al., 2016; Jonczak et al., 2023). En etapas más tempranas, sin embargo, dicha relación
podía ser positiva (Velázquez et al., 2023). El tipo de vegetación existente en los nuevos bosques y
zonas de matorral desarrollados en tierras agrarias abandonadas también
resultaba determinante para la acumulación de C en el suelo. Al estudiar la
fijación de C en el suelo de los nuevos bosques desarrollados en pastizales
abandonados de los Alpes y los Apeninos, Pellis et
al. (2019) detectaron un mayor COS en los
Apeninos, donde los bosques estaban dominados por especies de hoja caduca que,
en los Alpes, donde estaban dominados por especies de hoja perenne. En
ambientes mediterráneos, sin embargo, la acumulación del COS en tierras
agrarias abandonadas venía determinada por la cobertura de especies herbáceas (Gabarrón-Galeote et al., 2015).
Esto puede explicarse por el hecho de que, en este tipo de especies, la mayor
parte de los aportes de MO proceden de las raíces, por lo que están, por tanto,
más protegidos frente a la degradación (Meyer et
al., 2012) (Fig. A4
del Anexo).
Restauración, gestión forestal y territorial
La colonización vegetal de tierras
agrarias abandonadas ha resultado clave para el proceso de expansión forestal
que ha tenido lugar en el continente europeo (Palmero-Iniesta
et al., 2021). Los nuevos bosques y zonas de matorral desarrollados en
tierras agrarias abandonadas son una realidad, por lo que, durante la última
década ha cobrado fuerza la idea de que las estrategias dirigidas a una
correcta gestión de la regeneración natural de estas zonas resultan más
efectivas (y menos costosas) para mitigar los efectos del cambio climático que
las enfocadas al aumento (aún mayor) de la superficie forestal mediante la
plantación de árboles (Tölgyesi et al., 2022). El
desarrollo de ciertas actuaciones en estos nuevos bosques y zonas de matorral
resulta crucial, además, para favorecer su adaptación al cambio climático. Por
ejemplo, los hayedos desarrollados en tierras agrarias abandonadas de la
Cataluña interior muestran un mayor crecimiento y una menor densidad de la
madera que los hayedos maduros situados en zonas cercanas, lo que les hace ser
más sensibles a las sequías (Alfaro-Sánchez et
al., 2019). En este caso, la realización de cortas selectivas haría que los
árboles persistentes resistieran mejor este fenómeno (Sohn
et al., 2016). Asimismo, la realización de quemas prescritas en los bosques
desarrollados en tierras agrarias abandonadas puede contribuir a aumentar el
contenido de C mineral en el suelo (Nave et al., 2022).
En aquellas zonas donde el matorral desarrollado tras el abandono agrícola
muestre una mayor densidad pueden realizarse clareos, lo que provocaría un
aumento de la fijación de C en la vegetación restante, teniendo efectos
positivos para la regulación del ciclo hidrológico (Korchani
et al., 2022) y la prevención de incendios forestales (Madrigal et al., 2017). En general, las actuaciones en
el marco de una silvicultura moderada no sólo tendrían un efecto positivo en
las reservas de C existentes en el suelo (Bernués et
al., 2011) y la biomasa (Aldea et al., 2017) de
los nuevos bosques y zonas de matorral desarrollados en tierras agrarias
abandonadas, si no que, además, contribuirían a evitar la extracción ilegal de
madera y leña (Chiti et al., 2018).
Pese a todo lo expuesto líneas
arriba, es importante señalar que en las tierras agrarias abandonadas en las
que no existan fuentes de propágulos en las proximidades (e.g. bosques maduros,
setos vegetales o árboles aislados), la colonización vegetal va a ser lenta o
inexistente (Cuesta et al., 2012). Por otra parte, en aquellas tierras agrarias abandonadas
en las que exista una fuerte degradación del suelo (Robledano-Aymerich et al., 2014), la colonización vegetal no va a contribuir por si sola a
la reducción de la erosión (Rodrigo-Comino
et al., 2018; Cortijos-López
et al., 2023). En este tipo de áreas, por tanto,
puede ser necesario desarrollar estrategias de restauración activa (Selwyn et al., 2023). Estas
estrategias no consisten sólo en la plantación de árboles, sino también en la
implantación de posaderos para facilitar la dispersión de semillas, el
desarrollo de un pastoreo de baja intensidad para favorecer la diversidad de
especies, o la introducción de plantas leguminosas para aumentar la
disponibilidad de N y el secuestro de C en el suelo y la vegetación (Bonet, 2004). También es
importante desarrollar actuaciones para aumentar la cobertura de especies
herbáceas, a fin de promover la estabilización de las reservas de C en el suelo
(Thuille y Schulze, 2006; Gabarrón-Galeote et al.,
2015). Por último, en tierras agrarias abandonadas de suelos arenosos y con
un bajo contenido inicial en C, pueden realizarse enmiendas orgánicas a fin de
favorecer e aumento del COS (ej. Halassy et al., 2020).
Desde un punto de vista más relacionado con la gestión
territorial, y para la península ibérica, Lasanta et al.
(2024) y Lloret et al. (2024) han señalado
recientemente que la promoción de un uso extensivo del territorio sería la
estrategia más exitosa para favorecer un aumento de la fijación de C y la
biodiversidad de los ecosistemas. Dicha estrategia contribuiría, además, al
desarrollo de un paisaje más fragmentado, diverso y resiliente frente a
perturbaciones como los incendios forestales, así como a la lucha contra la
despoblación de las zonas de media montaña.
Conclusiones
1. La gran mayoría de los trabajos sobre fijación de C durante
la colonización vegetal en tierras agrarias abandonadas realizados en Europa se
han centrado en el análisis de la dinámica del C en el suelo, siendo muchos
menos los que han tenido en cuenta dinámica del C en otros compartimentos del
ecosistema, así como los flujos de C existentes entre compartimentos.
2. El
COS, en general, aumenta durante la colonización vegetal
de tierras agrarias abandonadas, pero sigue patrones muy variados relacionados
con el clima, el tipo de uso previo al abandono y los cambios en los aportes de
C tras la colonización vegetal de especies leñosas. En general, se acumula más
C en el suelo de aquellos sistemas que partían de un contenido inicial menor de
este elemento.
3. Durante
la sucesión secundaria en tierras agrarias abandonadas, el CBA sigue un claro
patrón de aumento, pero puede alcanzar un máximo a partir de 60-80 años después
del abandono, debido a la existencia de procesos de auto-aclareo.
4. Entre los
factores determinantes de la acumulación de C en el suelo durante el proceso de
colonización vegetal en tierras agrarias abandonadas destacan la PMA, que se
relaciona de forma positiva con el COS en el suelo superficial hasta un máximo
de 800-1100 mm, y la TMA que se relaciona de forma positiva con el COS en todo
el perfil. Ambas relaciones vienen explicadas por los cambios en la actividad
microbiana y la descomposición de la hojarasca en los distintos tipos de
climas.
5. Otros
factores determinantes de la acumulación de C en el suelo en tierras agrarias
abandonadas son el tipo de uso agrícola previo al abandono (se produce una
mayor acumulación de COS en antiguos cultivos leñosos), y el contenido en N,
que suele aumentar de forma paralela al COS.
6. Una
mayor densidad de individuos en los nuevos bosques desarrollados en tierras
agrarias abandonadas provoca un aumento en el CBA durante las etapas iniciales,
y una disminución en etapas posteriores, antes de que se produzcan los procesos
de auto-aclareo.
7. El
secuestro de C en el suelo y la biomasa de los nuevos bosques y matorrales
desarrollados en tierras agrarias abandonadas puede favorecerse llevando a cabo
cortas selectivas, quemas prescritas y clareos, e impulsando usos como la
ganadería extensiva en las zonas afectadas por el abandono agrícola.
Disponibilidad de datos y código
Los datos y el código de
R utilizados para la realización de los análisis del presente artículo se
encuentran disponibles en abierto, en el siguiente enlace:
https://zenodo.org/records/19144222?token=eyJhbGciOiJIUzUxMiJ9.eyJpZCI6ImY2OGQzOWJjLTFjMzQtNDc3Yy04ZTQyLWUxOTQzNDQwOTVlNyIsImRhdGEiOnt9LCJyYW5kb20iOiI4ZGJjMzE3ZTEzZjcwM2JjM2FkMzc3YTg3ODNmZmUxZiJ9.YRGvlHhb21tZ2Vl-1-BBT8IdYMn85Ab6qTb_F21QxXM4UGVAwL8gNEn37CVL6bd5eF1JckVVLFKGiiFrCDzIQQ
Financiación, permisos requeridos, potenciales
conflictos de interés y agradecimientos”
Este estudio ha sido financiado por la Comisión Europea a
través del proyecto CASE-CO2 (H2020-MSCA-IF-2017, DLV; 799885).
El autor declara no tener ningún conflicto de interés.
El autor quiere expresar su agradecimiento a A. Escudero y
J. Vayreda por sus sugerencias durante la preparación de la primera propuesta
del proyecto CASE-CO2, así como a F. Bravo y Mª. B. Turrión por su apoyo
durante el desarrollo del mismo. También quiere agradecer a J. Company, N.
Delcourt, J. Jonczak, M. B. Valjavec, I. Kurganova y Mart-Jan Schelhaas por los
datos, comentarios y fotografías aportados para el presente trabajo.
Referencias
Acín-Carrera, M., Marques, M.
J., Carral, P., Álvarez, A. M., López, C., Martín-López, B., & González, J.
A. (2013). Impacts of land-use intensity on soil organic
carbon content, soil structure and water-holding capacity. Soil Use and
Management, 29, 547–556. https://doi.org/10.1111/sum.12064
Acuña-Mínguez, B., Rincón, A., Valladares, F., Cordero, I., & Bravo-Oviedo,
A. (2024). Natural forest colonisation as a management strategy to restore soil
functioning of abandoned agricultural lands. Journal of Applied Ecology, 61,
2690–2700. https://doi.org/10.1111/1365-2664.14780
Alberti, G., Peressotti, A., Piussi, P., & Zerbi, G. (2008). Forest
ecosystem carbon accumulation during a secondary succession in the Eastern
Prealps of Italy. Forestry, 81, 1–11. https://doi.org/10.1093/forestry/cpm026
Alberti, G., Leronni, V., Piazzi, M., Petrella, F., Mairota, P.,
Peressotti, A., … Rühl, J. (2011). Impact of woody encroachment on soil organic
carbon and nitrogen in abandoned agricultural lands along a rainfall gradient
in Italy. Regional Environmental Change, 11, 917–924. https://doi.org/10.1007/s10113-011-0229-6
Aldea, J., Bravo, F., Bravo-Oviedo,
A., Ruiz-Peinado, R., Rodríguez, F., & Del Río, M. (2017). Thinning enhances the species-specific radial increment response to
drought in Mediterranean pine–oak stands. Agricultural and Forest
Meteorology, 237, 371–383. https://doi.org/10.1016/j.agrformet.2017.02.009
Alfaro-Sánchez, R., Jump, A. S., Pino, J., Díez-Nogales, O., & Espelta, J. M.
(2019). Land use legacies drive higher growth, lower wood density and enhanced
climatic sensitivity in recently established forests. Agricultural and
Forest Meteorology, 276–277, 107630. https://doi.org/10.1016/j.agrformet.2019.107630
Anderson, K. J., Allen, A. P., Gillooly, J. F., & Brown, J. H. (2006).
Temperature dependence of biomass accumulation rates during secondary
succession. Ecology Letters, 9, 673–682. https://doi.org/10.1111/j.1461-0248.2006.00914.x
Aosaar, J., Mander, Ü., Varik, M., Becker, H., Morozov, G., &
Maddison, M. (2016). Biomass production and nitrogen balance of naturally
afforested silver birch (Betula pendula Roth.) stand in Estonia. Silva
Fennica, 50, 1628. https://doi.org/10.14214/sf.1628
Badalamenti, E., Battipaglia,
G., Gristina, L., Novara, A., Rühl, J., Sala, G., … La Mantia,
T. (2019). Carbon stock increases up to old growth
forest along a secondary succession in Mediterranean island ecosystems. PLOS
ONE, 14, e0220194. https://doi.org/10.1371/journal.pone.0220194
Beck, H. E., McVicar, T. R.,
Vergopolan, N., Berg, A., Lutsko, N. J., Dufour, A., … Miralles,
D. (2023). High-resolution Köppen–Geiger maps for
1901–2099 based on constrained CMIP6 projections. Scientific
Data, 10, 724. https://www.nature.com/articles/s41597-023-02549-6
Beilin, R., Lindborg, R., Stenseke, M., Pereira, H. M., Llausàs, A.,
Slätmo, E., … Queiroz, C. (2014). Analysing how drivers
of agricultural land abandonment affect biodiversity and cultural landscapes
using case studies from Scandinavia, Iberia and Oceania. Land Use Policy, 36,
60–72. https://doi.org/10.1016/j.landusepol.2013.07.003
Bell, S. M., Terrer, C., Barriocanal, C., Jackson, R. B., &
Rosell-Melé, A. (2021). Soil organic carbon accumulation rates on Mediterranean
abandoned agricultural lands. Science of the Total Environment, 759,
143535. https://doi.org/10.1016/j.scitotenv.2020.143535
Bernués, A., Ruiz, R., Olaizola,
A., Villalba, D., & Casasús, I. (2011). Sustainability of
pasture-based livestock farming systems in the European Mediterranean context:
Synergies and trade-offs. Livestock Science, 139, 44–57. https://doi.org/10.1016/j.livsci.2011.03.018
Bijak, S., Zasada, M., Bronisz, A., Bronisz, K., Czajkowski, M.,
Ludwisiak, Ł., … Wojtan, R. (2013). Estimating coarse roots biomass in young
silver birch stands on post-agricultural lands in central Poland. Silva
Fennica, 47, Article 963. https://doi.org/10.14214/sf.963
Bohner, A., Karrer, J., Walcher, R., Brandl, D., Michel, K., Anrberger,
A., … Zaller, J. G. (2019). Ecological responses of semi-natural grasslands to
abandonment: Case studies in three mountain regions in the Eastern Alps. Folia
Geobotanica, 54, 211–225. https://doi.org/10.1007/s12224-019-09355-2
Bonet, A. (2004). Secondary succession of semi-arid Mediterranean
old-fields in south-eastern Spain: Insights for conservation and restoration of
degraded lands. Journal of Arid Environments, 56, 213–233. https://doi.org/10.1016/S0140-1963(03)00048-X
Bormann, B.
T., & Gordon, J. C. (1984). Stand density
effects in young red alder plantations: Productivity, photosynthate
partitioning and nitrogen fixation. Ecology, 65, 394–402.
Bose, A., Schelhaas, M.-J., Mazerolle, M. J., & Bongers, F. (2014). Temperate forest development during secondary succession: Effects of
soil, dominant species and management. European Journal of Forest
Research, 133, 511–523. https://doi.org/10.1007/s10342-014-0781-y
Calvo de Anta, R., Luís, E.,
Febrero-Bande, M., Galiñanes, J., Macías, F., Ortíz, R., & Casás, F.
(2020). Soil organic carbon in peninsular Spain: Influence of
environmental factors and spatial distribution. Geoderma,
370, Article 114365. https://doi.org/10.1016/j.geoderma.2020.114365
Castro, H., Fortunel, C., & Freitas, H. (2010). Effects of land abandonment on plant litter decomposition in a
Montado system: Relation to litter chemistry and community functional
parameters. Plant and Soil, 333, 181–190. https://doi.org/10.1007/s11104-010-0333-2
Chiti, T., Emanuele, B., Guido, P., Lucia, P., Chiriacò, M.V., &
Riccardo, V., (2018). Soil organic carbon pool's contribution to climate change
mitigation on marginal land of a Mediterranean montane area in Italy. Journal
of Environmental Management 218: 593-601. https://doi.org/10.1016/j.jenvman.2018.04.093
Clemmensen, K.E., Bahr, A., Ovaskainen, O., Dahlberg, A., Ekblad, A.,
Wallander, H., … Lindahl, B.D. (2013). Roots and associated fungi drive
long-term carbon sequestration in boreal forest. Science 339 :
1615–1618. https://doi.org/10.1126/science.1231923
Company, J., Valiente, N.,
Fortesa, J., García-Comendador, J., Lucas-Borja, M. E., Ortega, R., … Estrany, J. (2022). Secondary succession and
parent material drive soil bacterial community composition in terraced
abandoned olive groves from a Mediterranean hyper-humid mountainous area.
Agriculture, Ecosystems and Environment 332: 107932. https://doi.org/10.1016/j.agee.2022.107932
Cortez, J., Garnier, E., Pérez-Harguindeguy, N., Debussche, M., &
Gillon, D. (2007). Plant traits, litter quality and
decomposition in a Mediterranean old-field succession. Plant and Soil
296: 19-34. https://doi.org/10.1007/s11104-007-9285-6
Cortijos-López, M.,
Sánchez-Navarrete, P., Lasanta, T., & Nadal-Romero, E. (2023). How do acid or alkaline soil environments affect soil organic carbon
stocks in a post-abandonment secondary succession process in Mediterranean
mountain areas? Catena, 232, 107384. https://doi.org/10.1016/j.catena.2023.107384
Cramer, V. A., Hobbs, R. J., & Standish, R. J. (2008). What’s new
about old fields? Land abandonment and ecosystem assembly. Trends in Ecology
& Evolution, 23, 104–112. https://doi.org/10.1016/j.tree.2007.10.005
Crawley, M. J. (2007). The R book. John Wiley & Sons.
Cuesta, B., Rey Benayas, J. M.,
Gallardo, A., Villar-Salvador, P., & González-Espinosa, M. (2012). Soil chemical properties in abandoned Mediterranean cropland after
succession and oak reforestation. Acta Oecologica, 38, 58–65. https://doi.org/10.1016/j.actao.2011.09.004
Doblas-Miranda, E., Rovira,
P., Brotons, L., Martinez-Vilalta, J., Retana, J., Pla, M., & Vayreda, J.
(2013). Soil carbon stocks and their variability across the
forests, shrublands and grasslands of peninsular Spain. Biogeosciences,
10, 8353–8361. https://doi.org/10.5194/bg-10-8353-2013
Facioni, L., Burrascano, S.,
Chiti, T., Giarrizzo, E., Zanini, M., & Blasi, C. (2019). Changes
in plant diversity and carbon stocks along a succession from semi-natural
grassland to sub-Mediterranean Quercus cerris L. woodland in Central
Italy. Phytocoenologia, 49, 393–408. https://doi.org/10.1127/phyto/2019/0299
Fuchs, R., Schulp, C. E., Hengeveld, G. M., Verburg, P. H., Clevers, J.
G. P. W., Schelhaas, M.-J., & Herold, M. (2016). Assessing the influence of
historic net and gross land changes on the carbon fluxes of Europe. Global
Change Biology, 22, 2526–2539. https://doi.org/10.1111/gcb.13191
Gabarrón-Galeote, M.A., Trigalet,
S. & van Wesemael, B. (2015). Effect of land abandonment
on soil organic carbon fractions along a Mediterranean precipitation gradient. Geoderma
249-250, 69-78. http://dx.doi.org/10.1016/j.geoderma.2015.03.007
Gamper, S. M., Tasser, E., & Tappeiner, U. (2007). Short‑time effects
of land‑use changes on O‑horizon in subalpine grasslands. Plant and Soil,
299, 101–115. https://doi.org/10.1007/s11104-007-9366-6
Glenn-Lewin, D. C., Peet, R. K., & Veblen, T. T. (1992). Plant
succession: Theory and prediction (Population and Community Biology Series
11). Cambridge University Press.
Guidi, C., Magid, J., Rodeghiero, M., Gianelle, D., & Vesterdal, L.
(2014). Effects of forest expansion on mountain grassland: Changes within soil
organic carbon fractions. Plant and Soil, 385, 373–387. https://doi.org/10.1007/s11104-014-2315-2
Haddaway, N. R., Styles, D., & Pullin, A. S. (2014). Environmental
impacts of farmland abandonment in high altitude/mountain regions: A systematic
map of the evidence. Environmental Evidence, 3, 17. https://doi.org/10.1186/2047-2382-3-17
Halassy, M., Kövendi-Jakó, A.,
Reis, B. P., Szitár, K., Seyidova, Z., & Török, K. (2020). N immobilization treatment revisited: A retarded and temporary
effect unfolded in old-field restoration. Applied Vegetation Science, 24,
e12555. https://doi.org/10.1111/avsc.12555
Heinman,
H., & Reichstein, M. (2008). Terrestrial
ecosystem carbon dynamics and climate feedbacks. Nature, 451, 289–292. https://doi.org/10.1038/nature06591
Herrero, C., Krankina, O., Monleón, V. J., & Bravo, F. (2014). Amount
and distribution of coarse woody debris in pine ecosystems of North‑Western
Spain, Russia and the United States. iForest, 7, 53–60. https://doi.org/10.3832/ifor0644-007
Hooker, T. D.,
& Compton, J. E. (2003). Forest ecosystem
carbon and nitrogen accumulation during the first century after agricultural
abandonment. Ecological Applications, 13, 299–313.
Ibáñez, I., Katz, D. S. W., Peltier, D., Wolf, S. M., & Connor Barrie,
B. T. (2014). Assessing the integrated effects of landscape fragmentation on
plants and plant communities: The challenge of multiprocess–multiresponse
dynamics. Journal of Ecology, 102, 882–895. https://doi.org/10.1111/1365-2745.12223
Jackson, R. B., Banner, J. L., Jobbágy, E. G., Pockman, W. T., & Wall,
D. H. (2002). Ecosystem carbon loss with woody plant invasion of grasslands. Nature,
418, 623–626. https://doi.org/10.1038/nature00910
Johnson, C., Zarin, D., & Johnson, A. (2000). Post-disturbance
aboveground biomass accumulation in global secondary forests. Ecology, 81,
1395–1401. https://doi.org/10.2307/177216
Jonczak, J., Oktaba, L., Pawlowicz, E., Chojnacka, A., Regulska, E.,
Słowinska, S., … Wójcik‑Gront, E. (2023). Soil organic matter transformation
influenced by silver birch (Betula pendula Roth) succession on abandoned
from agricultural production sandy soil. European Journal of Forest
Research, 142, 367–379. https://doi.org/10.1007/s10342-022-01527-8
Kalinina, O., Goryachkin, S. V., Lyuri, D. I., & Giani, L. (2015). Post‑agrogenic
development of vegetation, soils, and carbon stocks under self‑restoration in
different climatic zones of European Russia. Catena, 129, 18–29. https://doi.org/10.1016/j.catena.2015.02.016
Kalinina, O., Chertiv, O., Frolov, P., Goryachkin, S., Kuner, P., Küper, J.,
… Giani, L. (2018). Alteration process during the post‑agricultural restoration
of Luvisols of the temperate broad‑leaved forest in Russia. Catena, 171,
602–612. https://doi.org/10.1016/j.catena.2018.08.004
Kauppi, P. E., Ausubel, J. H., Fang, J., Mather, A. S., Sedjo, R. A.,
& Waggoner, P. E. (2006). Returning forests analyzed with the forest
identity. Proceedings of the National Academy of Sciences, 103, 47. https://doi.org/10.1073/pnas.0608343103
Knops, J. M.
H., & Tilman, D. (2000). Dynamics of soil
nitrogen and carbon accumulation for 61 years after agricultural abandonment. Ecology,
81, 88–98.
Kopecký,
M., & Vojta, J. (2009). Land‑use legacies in
post‑agricultural forests in the Doupovské Mountains, Czech Republic. Applied
Vegetation Science, 12, 251–260. https://doi.org/10.1111/j.1654-109X.2009.01023.x
Korchani, M., Nadal‑Romero, E., Lasanta, T., & Tague, C. (2022). Carbon
sequestration and water yield trade‑offs following restoration of abandoned
agricultural lands in Mediterranean mountains. Environmental
Research, 207, 112203. https://doi.org/10.1016/j.envres.2021.112203
Kurganova, I., Lopes de
Guerenyu, V., & Kuzyakov, Y. (2015). Large‑scale carbon
sequestration in post‑agrogenic ecosystems in Russia and Kazakhstan. Catena,
133, 461–466. https://doi.org/10.1016/j.catena.2015.06.002
Kurganova, I., Merino, A., Lopes
de Gerenyu, V., Barros, N., Kalinina, O., Giani, L. & Kusyakov, Y. (2019). Mechanisms of carbon sequestration and stabilization by restoration
of arable soils after abandonment: A chronosequence study on Phaeozems and
Chernozems. Geoderma 354, 113882. https://doi.org/10.1016/j.geoderma.2019.113882
Laganière, J., Angers, D. A., & Paré, D. (2010). Carbon accumulation in
agricultural soils after afforestation: A meta‑analysis. Global
Change Biology, 16, 439–453. https://doi.org/10.1111/j.1365-2486.2009.01930.x
La Mantia, T., Gristina, L.,
Rivaldo, E., Pasta, S., Novara, A. & Rühl, J. (2013). The
effects of post-pasture woody plant colonization on soil and aboveground litter
carbon and nitrogen along a bioclimatic transect. iForest 6 (4),
238–246. https://doi.org/10.3832/ifor0811-006
Lal, R. (2004). Soil carbon sequestration to mitigate climate change. Geoderma,
123, 1–22. https://doi.org/10.1016/j.geoderma.2004.01.032
Lasanta, T., Sánchez‑Navarrete,
P., Medrano‑Moreno, L. M., Khorchani, M., & Nadal‑Romero, E. (2020). Soil quality and soil organic carbon storage in abandoned
agricultural lands: Effects of revegetation processes in a Mediterranean mid‑mountain
area. Land Degradation & Development, 31, 2830–2845. https://doi.org/10.1002/ldr.3655
Lasanta, T., Cortijos-López, M.,
Errea, M. P., Llena, M., Sánchez-Navarrete, P., Zabalza, J., &
Nadal-Romero, E. (2024). Shrub clearing and extensive
livestock as a strategy for enhancing ecosystem services in degraded
Mediterranean mid-mountain areas. Science of the Total Environment, 906,
167668. https://doi.org/10.1016/j.scitotenv.2023.167668
Law, B. E., Sun, O. J., Campbell, J., van Tuyl, S., & Thornton, P.
E. (2003). Changes in carbon storage and fluxes in a chronosequence of
ponderosa pine. Global Change Biology, 9, 510–524. https://doi.org/10.1046/j.1365-2486.2003.00624.x
Lichner, L., Sŭrda, P., Toková, L., Holos, S., Kollár, J., & Igaz, D.
(2023). Impact of duration of land abandonment on soil properties. Journal
of Hydrology and Hydromechanics, 71, 148–155. https://doi.org/10.2478/johh-2023-0011
Lloret, F., Escudero, A., Lloret,
J., & Valladares, F. (2024). An ecological perspective for
analysing rural depopulation and abandonment. People and Nature, 6,
490–506. https://doi.org/10.1002/pan3.10606
Luyssaert, S., Schulze, E.-D., Börner, A., Knohl, A., Hessenmöller, D., Law,
B. E., Ciais, P., & Grace, J. (2008). Old-growth forests as global carbon
sinks. Nature, 455, 213–215. https://doi.org/10.1038/nature07276
MacDonald, D., Crabtree, J. R., Wiesinger, G., Dax, T., Stamou, N., Fleury,
P.,… Gibon, A. (2000). Agricultural abandonment in mountain areas of Europe:
Environmental consequences and policy response. Journal of
Environmental Management, 59, 47–69.
Madrigal, J.,
Fernández-Migueláñez, I., Hernando, C., Guijarro, M., Vega-Nieva, D. J., &
Tolosana, E. (2017). Does forest biomass harvesting for energy
reduce fire hazard in the Mediterranean basin? A case study in the Caroig
Massif (Eastern Spain). European Journal of Forest Research, 136, 13–26.
https://doi.org/10.1007/s10342-016-1004-5
Marzaioli, R., D’Ascoli, R., De
Pascale, R. A., & Rutigliano, F. A. (2010). Soil quality
in a Mediterranean area of Southern Italy as related to different land-use
types. Applied Soil Ecology, 44, 205–212. https://doi.org/10.1016/j.apsoil.2009.12.007
McLauchlan, K. (2006). The nature and longevity of agricultural impacts on
soil carbon and nutrients: A review. Ecosystems, 9, 1364–1382.
Meyer, S., Leifeld, J., Bahn, M., & Fuhrer, L. (2012). Free and
protected soil organic carbon dynamics respond differently to abandonment of
mountain grassland. Biogeosciences, 9, 853–865. https://doi.org/10.5194/bg-9-853-2012
Molina, J. A., Martín-Sanz, P. M.,
Valverde-Asenjo, I., Sánchez-Jiménez, A., & Quintana, J. R. (2023). Mediterranean grassland succession as an indicator of changes in
ecosystem biodiversity and functionality. Biodiversity and Conservation, 32,
95–118. https://doi.org/10.1007/s10531-022-02481-y
Moreno, G., Gallardo, J. F., & Vicente, M. A. (2011). How
Mediterranean deciduous trees cope with long summer drought? The case of Quercus
pyrenaica forests in Western Spain. In Bredemeier, M., Cohen, S., Godbold,
D. L., Lode, E., Pichler, V., & Schleppi, P. (Eds.), Forest management
and the water cycle: An ecosystem-based approach (Ecological Studies 212,
pp. 189–201). Springer-Verlag.
Nadal-Romero, E., Cammeraat,
E., Pérez-Cardiel, E., & Lasanta, T. (2016). How do soil
organic carbon stocks change after cropland abandonment in Mediterranean humid
mountain areas? Science of the Total Environment, 566–567, 741–752. https://doi.org/10.1016/j.scitotenv.2016.05.031
Nadal-Romero, E., Otal-Laín,
I., Lasanta, T., Sánchez-Navarrete, P., Errea, P., & Cammeraat, E. (2018). Woody encroachment and soil carbon stocks in subalpine areas in the
Central Spanish Pyrenees. Science of the Total Environment, 636,
727–736. https://doi.org/10.1016/j.scitotenv.2018.04.324
Navas, A., Gaspar, L., Quijani, L.,
López-Vicente, M., & Machín, J. (2012). Patterns of soil
organic carbon and nitrogen in relation to soil movement under different land
uses in mountain fields (South Central Pyrenees). Catena, 94, 43–52. https://doi.org/10.1016/j.catena.2011.05.012
Nave, L. E., DeLyser, K., Domke, G. M., Holub, S. M., Janowiak, M. K.,
Ontl, T. A., … Swanston, C. W. (2022). Soil carbon in the South Atlantic United
States: Land use change, forest management, and physiographic context. Forest
Ecology and Management, 520, 120410. https://doi.org/10.1016/j.foreco.2022.120410
Nóvak, T. J., Incze, J., Spohn, M., Glina, B., & Giani, L. (2014).
Soil and vegetation transformation in abandoned vineyards of the Tokaj
Nagy-Hill, Hungary. Catena, 123, 88–98. https://doi.org/10.1016/j.catena.2014.07.017
Novara, A., Gristina, L., Sala, G.,
Galati, A., Crescimanno, M., Cerdà, A., … La Mantia, T.
(2017). Agricultural land abandonment in Mediterranean
environment provides ecosystem services via soil carbon sequestration. Science
of the Total Environment, 576, 420–429. https://doi.org/10.1016/j.scitotenv.2016.10.123
Oddi, L., Bergò, S. E., Lerda, C.,
Bresciani, L., Galvagno, M., Celi, L. R., & Siniscalco, C. (2025). Consequences of ten-year abandonment in a subalpine grassland: Soil
and litter decomposition changes. Botanical Journal of the Linnean Society,
4, 424–434. https://doi.org/10.1093/botlinnean/boaf018
Ovsepyan, L., Kurganova, I.,
Lopes de Guerenyu, V., & Kuzyakov, Y. (2019). Recovery of
organic matter and microbial biomass after abandonment of degraded agricultural
soils: The influence of climate. Land Degradation & Development, 30,
1861–1874. https://doi.org/10.1002/ldr.3387
Palmero-Iniesta, M., Pino, J., Pesquer, L., & Espelta, J. M. (2021). Recent
forest area increase in Europe: Expanding and regenerating forests differ in
their regional patterns, drivers and productivity trends. European Journal
of Forest Research, 140, 793–805. https://doi.org/10.1007/s10342-021-01366-z
Palombo, C., Chirici, G., Marchetti, M., & Tognetti, R. (2013). Is land
abandonment affecting forest dynamics at high elevation in Mediterranean
mountains more than climate change? Plant Biosystems, 147, 1–11. https://doi.org/10.1080/11263504.2013.772081
Peel, M. C., Finlayson, B. L., & McMahon, T. A. (2007). Updated
world map of the Köppen–Geiger climate classification. Hydrology and Earth
System Sciences, 11, 1633–1644. https://doi.org/10.5194/hess-11-1633-2007
Pellis, G., Chiti, T., Rey, A., Curiel-Yuste, J., Trotta, C., &
Papale, D. (2019). The ecosystem carbon sink implications of mountain forest
expansion into abandoned grazing land: The role of subsoil and climatic
factors. Science of the Total Environment, 672, 106–120. https://doi.org/10.1016/j.scitotenv.2019.03.329
Peet, R. K. (1981). Changes in biomass and production during secondary
forest succession. In West, D. C., Shugart, H. H., & Botkin, D. B. (Eds.), Forest
succession: Concepts and applications (pp. 324–338). Springer.
Poeplau, C., Don, A., Vesterdal, L., Leifeld, J., van Wesemael, B.,
Schumacher, J., & Gensior, A. (2011). Temporal dynamics of soil organic
carbon after land-use change in the temperate zone: Carbon response functions
as model approach. Global Change Biology, 17, 2415–2427. https://doi.org/10.1111/j.1365-2486.2011.02408.x
Post, W. M.,
& Kwon, K. C. (2000). Soil carbon sequestration
and land-use change: Processes and potential. Global Change Biology, 6,
317–327.
Poulton, P. R., Pye, E., Hargreaves, P. R., & Jenkinson, D. S. (2003).
Accumulation of carbon and nitrogen by old arable land reverting to woodland. Global
Change Biology, 9, 942–955. https://doi.org/10.1046/j.1365-2486.2003.00633.x
Ramankutty, N. (2006). Global land-cover change: Recent progress, remaining
challenges. In E. F. Lambin & H. J. Geist (Eds.), Land-use and
land-cover change (pp. 9–41). Springer-Verlag.
Rastetter, E. B., McKane, R. B., Shaver, G. R., & Melillo, J. M. (1992).
Changes in C storage by terrestrial ecosystems: How C:N interactions restrict
responses to CO₂ and temperature. Water, Air, and Soil Pollution, 64,
327–344.
Rhemtulla, J. M., Mladenoff, D. J., & Clayton, M. K. (2009). Historical
forest baselines reveal potential for continued carbon sequestration. Proceedings
of the National Academy of Sciences of the U.S.A., 106, 6082–6087. https://doi.org/10.1073/pnas.0810076106
Rhyzova, I. M., Telesnina, V. M.,
& Sitnikova, A. A. (2020). Dynamics of soil properties and
carbon stocks structure in postagrogenic ecosystems of Southern Taiga during
natural reforestation. Eurasian Soil Science, 53, 240–252. https://doi.org/10.1134/S1064229320020106
Risch, A., Jurgensen, M. F., Page-Dumroese, D. S., & Schütz, M.
(2008). Long-term development of above- and below-ground carbon stocks
following land-use change in subalpine ecosystems of the Swiss National Park. Canadian
Journal of Forest Research, 38, 1590–1602. https://doi.org/10.1139/X08-014
Robledano-Aymerich, F.,
Romero-Díaz, A., Belmonte-Serrato, F., Zapata-Pérez, V. M., Martínez-Hernández,
C., & Martínez-López, V. (2014). Ecogeomorphological
consequences of land abandonment in semiarid Mediterranean areas: Integrated
assessment of physical evolution and biodiversity. Agriculture,
Ecosystems & Environment, 197, 222–242. https://doi.org/10.1016/J.AGEE.2014.08.006
Rodrigo-Comino, J.,
Martínez-Hernández, C., Iserloh, T., & Cerdà, A. (2018). Contrasted
impact of land abandonment on soil erosion in Mediterranean agriculture fields.
Pedosphere, 28, 617–631. https://doi.org/10.1016/S1002-0160(17)60441-7
Romero-Díaz, A., Ruiz-Sinoga,
J. D., Robledano-Aymerich, F., Brevik, E. C., & Cerdà, A. (2017). Ecosystem responses to land abandonment in Western Mediterranean
Mountains. Catena, 149, 824–835. https://doi.org/10.1016/j.catena.2016.08.013
Ruecker, G., Schad, P., Alcubilla, M. M., & Ferrer, C. (1999). Natural
regeneration of degraded soils and site changes on abandoned agricultural
terraces in Mediterranean Spain. Land Degradation & Development, 9,
179–188.
Ruiz-Peinado, R., Montero, G., & del Río, M. (2012). Biomass models to
estimate carbon stocks for hardwood tree species. Forest Systems, 21,
42–52. https://doi.org/10.5424/fs/2112211-02193
Ruskule, A., Nikodemus, O., Kasparinska, Z., Kasparinskis, R., &
Brūmelis, G. (2012). Patterns of afforestation on abandoned agriculture land in
Latvia. Agroforestry Systems, 85, 215–231. https://doi.org/10.1007/s10457-012-9495-7
Schneider, M. V., Oberholzer, S., & Speranza, C. I. (2024). Revegetation
is key for soil organic carbon sequestration on abandoned and degraded land in
northern Spain. Geoderma Regional, 38, e00835. https://doi.org/10.1016/j.geodrs.2024.e00835
Selwyn, M., Pino, J., &
Espelta, J. M. (2023). Disentangling the importance of
intrinsic and extrinsic seed dispersal factors in forest restoration success: A
global review. Restoration Ecology, 31, e13868. https://doi.org/10.1111/rec.13868
Schierhorn, F., Müller, D., Beringer, T., Prishchepov, A. V., Kümmerle, T.,
& Balmann, A. (2013). Post-Soviet cropland abandonment and carbon
sequestration in European Russia, Ukraine and Belarus. Global Biogeochemical
Cycles, 27, 1175–1185. https://doi.org/10.1002/2013GB004654
Shi, S. W., Zhang, W., Zhang, P., Yu, Y., & Ying, F. (2013). A
synthesis of change in deep soil organic carbon stores with afforestation of
agricultural soils. Forest Ecology and Management, 296, 53–63. https://doi.org/10.1016/j.foreco.2013.01.026
Sohn, J. A., Saha, S., & Bauhus, J. (2016). Potential of forest
thinning to mitigate drought stress: A meta-analysis. Forest Ecology and
Management, 380, 261–273. https://doi.org/10.1016/j.foreco.2016.07.046
Sokolowska, J., Jósefowska, A., Wóznica, K., & Zaleski, T. (2020).
Succession from meadow to mature forest: Impacts on soil biological, chemical
and physical properties—Evidence from the Pieniny Mountains, Poland. Catena,
189, 104503. https://doi.org/10.1016/j.catena.2020.104503
Spohn, M., Nóvak, T. J., Incze, J., & Giani, L. (2016). Dynamics of
soil carbon, nitrogen, and phosphorus in calcareous soils after land-use
abandonment – A chronosequence study. Plant & Soil, 401, 185–196. https://doi.org/10.1007/s11104-015-2513-6
Stewart, C. E., Paustian, K., Conant, R. T., Plante, A. F., & Six, J.
(2007). Soil carbon saturation: Concept, evidence and evaluation. Biogeochemistry,
86, 19–31. https://doi.org/10.1007/s10533-007-9140-0
Thuille, A.,
& Schulze, E.-D. (2006). Carbon dynamics in
successional and afforested spruce stands in Thuringia and the Alps. Global
Change Biology, 12, 325–342. https://doi.org/10.1111/j.1365-2486.2005.01078.x
Thuille, A., Buchmann, N., & Schulze, E.-D. (2000). Carbon stocks and
soil respiration rates during deforestation, grassland use and subsequent
Norway spruce afforestation in the Southern Alps, Italy. Tree Physiology, 20,
849–857.
Tölgyesi, C., Buisson, E., Helm, A., Temperton, V. M., & Török, P.
(2022). Urgent need for updating the slogan of global climate actions from
“tree planting” to “restore native vegetation”. Restoration Ecology,
30, e13594. https://doi.org/10.1111/rec.13594
Trigalet, S., Gabarrón-Galeote,
M. A., van Oost, K., & van Wesemael, B. (2016). Changes in
soil organic carbon pools along a chronosequence of land abandonment in
southern Spain. Geoderma, 268, 14–21. https://doi.org/10.1016/j.geoderma.2016.01.014
Uri, V., Varik, M., Aosaar, J., Kanal, A., Kukumägi, M., & Lõhmus,
K. (2012). Biomass production and carbon sequestration in a fertile silver
birch (Betula pendula Roth) forest chronosequence. Forest Ecology and
Management, 267, 117–126. https://doi.org/10.1016/j.foreco.2011.11.033
Vaccari, F. P., Lugato, E., Gioli, B., D’Acqui, L., Genesio, L., Toscano,
P., … Miglietta, F. (2012). Land use change and soil organic carbon dynamics in
Mediterranean agro-ecosystems: The case study of Pianosa Island. Geoderma,
175–176, 29–36. https://doi.org/10.1016/j.geoderma.2012.01.021
Valjavec, M. B., Čarni, A., Žlindra, D., Zorn, M., & Marinšek, A. (2022).
Soil organic carbon stock capacity in karst dolines under different land uses. Catena,
218, 106548. https://doi.org/10.1016/j.catena.2022.106548
Van Hall, R. L., Cammeraat, L. H., Keesstra, S. D., & Zorn, M. (2017).
Impact of secondary vegetation succession on soil quality in a humid
Mediterranean landscape. Catena, 149, 836–843. https://doi.org/10.1016/j.catena.2016.05.021
Vargas, R., Allen, M.F. & Allen, E.B. (2008). Biomass and carbon
accumulation in a fire chronosequence of a seasonally dry tropical forest.
Global Change Biology, 14, 109–124. https://doi.org/10.1111/j.1365-2486.2007.01462.x
Velázquez, E., Martínez-Jaraíz, C., Wheeler, C., Mitchard, E.T.A. &
Bravo, F. (2022). Forest expansion in abandoned
agricultural lands has limited effect to offset carbon emissions from
Central-North Spain. Regional Environmental Change, 22, 132.
https://doi.org/10.1007/s10113-022-01978-0
Velázquez, E., Turrión, M. B.,
& Bravo, F. (2023). Carbon accumulation over 70 years of
old-field succession driven by live biomass in a Mediterranean continental
environment. Forest Ecology and Management, 548, 121422. https://doi.org/10.1016/j.foreco.2023.121422
Vilà-Cabrera, A., Espelta, J. M., Vayreda, J., & Pino, J. (2017). “New
forests” from the twentieth century are a relevant contribution for C storage
in the Iberian Peninsula. Ecosystems, 20, 130–143. https://doi.org/10.1007/s10021-016-0019-6
Walker, L. R., Wardle, D. A., Bardgett, R. D., & Clarkson, B. D.
(2010). The use of chronosequences in studies of ecological succession and soil
development. Journal of Ecology, 98, 725–736. https://doi.org/10.1111/j.1365-2745.2010.01664.x
Wang, H., Yue, C., Mao, Q., Zhao, J., Ciais, P., Li, W., … Mu, X.
(2020). Vegetation and species impacts on soil organic carbon sequestration
following ecological restoration over the Loess Plateau, China. Geoderma,
371, Article 114389. https://doi.org/10.1016/j.geoderma.2020.114389
Weissgeber, M., Chanteloup, L., & Bonis, A. (2024). Carbon stock increase
during post‑agricultural succession in central France: No change of the
superficial soil stock and high variability within forest stages. New
Forests, 55, 1533–1555. https://doi.org/10.1007/s11056-024-10044-y
Yang, Y. H., Luo, Y. Q., & Finzi, A. C. (2011). Carbon and nitrogen
dynamics during forest stand development: A global synthesis. New
Phytologist, 190, 977–989. https://doi.org/10.1111/j.1469-8137.2011.03645.x
Zeglin, L. H., Bottomley, P. J., Jumpponen, A., Rice, C. W., Arango, M.,
Lindsley, A., … Myrold, D. D. (2013). Altered precipitation regime affects the
function and composition of soil microbial communities on multiple time scales.
Ecology, 94, 2334–2345.
Zhou, H., Wang, J., Sun, M., Wang, J., & Yang, K. (2024). Evolution
of farmland abandonment research from 1993 to 2023 using CiteSpace‑based
scientometric analysis. Journal of Geovisualization and Spatial Analysis, 8,
20. https://doi.org/10.1007/s41651-024-00177-1
Zornoza, R., Guerrero, C.,
Mataix-Solera, J., Scow, K. M., Arcenegui, V., & Mataix‑Beneyto, J. (2009).
Changes in soil microbial community structure following the
abandonment of agricultural terraces in mountainous areas of Eastern Spain. Applied
Soil Ecology, 42, 315–323. https://doi.org/10.1016/j.apsoil.2009.05.011
Anexo / Annex
Tabla A1. Modelos lineales y no lineales
utilizados en las regresiones entre los valores máximos del carbono orgánico
del suelo (COS; Mg C ∙
ha-1) en el suelo superficial y en todo el perfil (0 ≤15 y 0 ≤ 80 cm de profundidad, respectivamente), la
temperatura y la precipitación media anual (TMA y PMA respectivamente), en los
sitios de estudio de 68 artículos científicos publicados entre 1996 y 2024 y
desarrollados en Europa. Los mismos modelos fueron utilizados en las
regresiones entre los valores máximos del carbono en la biomasa aérea (CBA; Mg C ∙
ha-1), la TMA y la PMA en los sitios de estudio de 13 artículos
científicos publicados durante el mismo periodo y desarrollados en el mismo
ámbito geográfico. En las ecuaciones, la letra Y hace referencia a los valores
máximos del COS y el CBA y la letra X a los valores de la TMA y la PMA.
Table A1. Linear and nonlinear models used in regressions
of maximum soil organic carbon values (COS; Mg C ∙ ha-1) in the
surface soil and throughout the profile (0 ≤ 15 and 0 ≤ 80 cm depth,
respectively), temperature, and annual average precipitation (TMA and PMA,
respectively), at the study sites of 68 scientific articles published between
1996 and 2024 and conducted in Europe. The same models were used in the
regressions between the maximum values of carbon in aboveground biomass (CBA;
Mg C ∙ ha-1), AAT, and AAP at the study sites of 13 scientific
articles published during the same period and conducted in the same geographic
area. In the equations, the letter Y refers to the maximum values of COS and
CBA, and the letter X refers to the values of TMA and PMA.
|
Modelo de regresión
|
Ecuación
|
|
(1)
Lineal
|
Y
~ a + b (X)
|
|
(3)
Asintótico de dos parámetros
|
Y
~ a × (1- exp(-b×X))
|
|
(2) Asintótico de tres parámetros
|
Y
~ a – b × exp(-c×X)
|
|
(4)
Michaelis-Menten
|
Y
~ (a × X) / (b+X)
|
Tabla A2.
Modelos de regresión entre los valores máximos del carbono orgánico del
suelo (COS; Mg C ∙ ha-1) en el suelo superficial y en todo el perfil
(0 ≤15 y 0 ≤ 80 cm de profundidad, respectivamente), la temperatura y la
precipitación media anual (TMA y PMA respectivamente), en los sitios de estudio
de 68 artículos científicos publicados entre 1996 y 2024, en Europa. Se indican los valores del criterio de información de
Akaike (AIC), la variación del criterio de información de Akaike (AIC), el
error medio residual cuadrado (RMSE), el valor y el error estándar (EE) de los
parámetros del modelo, y el coeficiente de determinación (R2) de los
modelos lineales. ** y * indican significación con P ≤ 0.001 y ≤ 0.05,
respectivamente. El modelo elegido (aquel con los valores más bajos de AIC y
RMSE), aparece señalado en negrita.
Table A2. Regression
models for the relationship between maximum soil organic carbon values (SOC; Mg
C ∙ ha-1) in the surface soil and throughout the profile (0 ≤ 15 and
0 ≤ 80 cm depth, respectively), mean annual temperature and precipitation (MAT
and MAP respectively), at the study sites of 68 scientific articles published
between 1996 and 2024 in Europe. The values of the Akaike information criterion
(AIC), the variation in the Akaike information criterion (AIC), the root mean
square error (RMSE), the value and standard error (SE) of the model parameters,
and the coefficient of determination (R²) of the linear models are shown. **
and * indicate significance at P ≤ 0.001 and ≤ 0.05, respectively. The chosen
model (the one with the lowest AIC and RMSE values) is highlighted in bold.
Tabla A3.
Modelos de regresión entre los valores máximos del carbono en la biomasa
aérea (CBA; Mg C ∙ ha-1), la TMA y la PMA en los sitios de estudio
de 13 artículos científicos publicados entre 1996 y 2024, en Europa. Se indican
los valores del criterio de información de Akaike (AIC), la variación del
criterio de información de Akaike (AIC), el error medio residual cuadrado
(RMSE), el valor y el error estándar (EE) de los parámetros del modelo, y el
coeficiente de determinación (R2) de los modelos lineales. El modelo
elegido (aquel con los valores más bajos de AIC y RMSE), aparece señalado en
negrita.
Table A3. Regression models relating the maximum values of
carbon in aboveground biomass (CBA; Mg C ∙ ha⁻¹), MAT and MAP
at the study sites of 13 scientific articles published between 1996 and 2024 in
Europe. The values of the Akaike information criterion (AIC), the variation in
the Akaike information criterion (AIC), the root mean square error (RMSE), the
value and standard error (SE) of the model parameters, and the coefficient of
determination (R²) of the linear models are shown. The chosen
model (the one with the lowest AIC and RMSE values) is highlighted in bold.

Figura A1. Toma de
muestras de suelo superficial en olivares abandonados de la Serra Tramuntana
(Mallorca, España). El suelo ha sido el compartimento del ecosistema más
estudiado en términos de fijación de C durante la sucesión secundaria en
tierras agrarias abandonadas en Europa. Aunque el suelo superficial (0-15 cm de
profundidad) es más activo secuestrando C, la mayor parte de las reservas de
este elemento se acumulan en el suelo profundo (> 15 cm de profundidad). Foto: Jaume Company.
Figure A1. Sampling of surface soil in abandoned olive
groves in the Serra Tramuntana (Mallorca, Spain). Soil has been the most
studied ecosystem compartment in terms of carbon sequestration during secondary
succession on abandoned agricultural land in Europe. Although the topsoil (0–15
cm deep) is more active in sequestering carbon, most of the carbon reserves
accumulate in the subsoil (> 15 cm deep). Photo:
Jaume Company.

Figura A2. Encinas (Quercus
ilex L.) colonizando un antiguo campo de centeno abandonado en Calzada de
Tera (Zamora, España). La colonización vegetal de plantas leñosas en tierras
agrarias abandonadas muestra un efecto de retraso (lag-effect), y suele tener
lugar entre 15-20 años después del abandono. Foto: el autor.
Figure A2. Holm oaks (Quercus ilex L.) colonising a former abandoned rye field in Calzada de
Tera (Zamora, Spain). The
colonisation of abandoned agricultural land by woody plants exhibits a lag
effect and usually takes place 15–20 years after abandonment. Photo: the author.

Figura
A3. Aspecto a
principios de primavera de un bosque dominado por Quercus robur L.
desarrollado en un campo de cultivo abandonado en la Reserva Forestal Kifhoek
(Países Bajos). Los suelos de textura fina, con una elevada proporción de limos
y arcillas, favorecen la acumulación de C en el suelo y, de forma indirecta, en
la biomasa. Foto: Mart-Jan Schelhaas.
Figure A3. A view in early spring of a forest dominated by Quercus robur L., which has developed on abandoned farmland in the Kifhoek
Forest Reserve (Netherlands). The fine-textured soils, with a high proportion
of silt and clay, promote carbon accumulation in the soil and, indirectly, in
the biomass. Photo: Mart-Jan Schelhaas.

Figura A4. Pastizal dominado por Lupinus
sp. desarrollado en un antiguo campo de cultivo en el Oblast Vladimir (Rusia).
Las herbáceas aportan una gran cantidad de residuos vegetales al suelo
superficial procedentes sobretodo de sus raíces, lo que contribuye a aumentar
el contenido en materia orgánica y, por ende, la acumulación de C. Foto: www.pikarabu.ru.
Figure A4. Grassland dominated by Lupinus sp., developed on a former arable field in the Vladimir
Oblast (Russia). The herbaceous plants contribute a large amount of plant
debris to the topsoil, mainly from their roots, which helps to increase the
organic matter content and, consequently, carbon sequestration. Photo: www.pikarabu.ru.