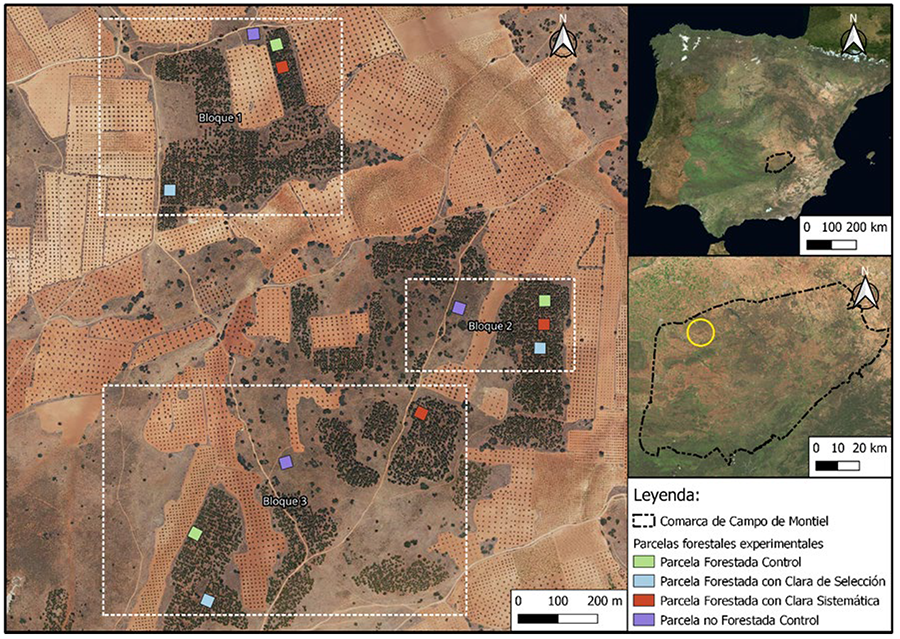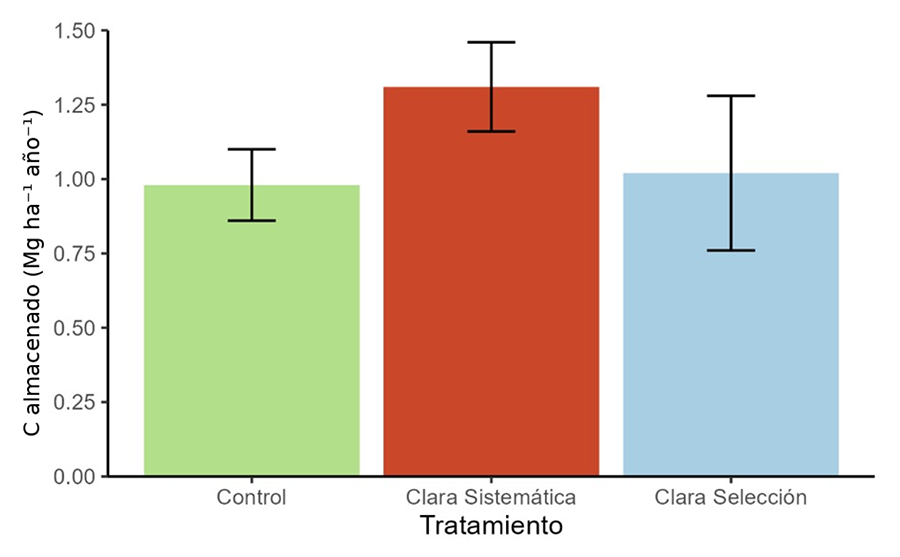
Figura 2. Tasa de carbono almacenado en la
biomasa durante el experimento. Las barras indican el promedio de las tres
parcelas con cada tratamiento ± desviación estándar. El carbono almacenado en
cada parcela se ha obtenido a partir de la suma de todos los pinos
individuales. No hay diferencias significativas entre los dos tipos de clara (Tabla A3 del Anexo).
Figure 2.
Carbon stocking rate during the experiment. Bars indicate the mean of the three
plots for each treatment ± standard deviation. The carbon stocked in each plot
was obtained by adding all individual pines. There are no significant
differences between the two thinning types (Table A3 del
Anexo).
Tabla 2. Biomasa
aérea, radical y total y carbono total almacenado durante el experimento. Las
cifras reportadas son el promedio de las tres parcelas con cada tratamiento ±
desviación estándar. La biomasa o el carbono de cada parcela se ha obtenido a
partir de la suma de todos los pinos individuales.
Table 2. Aerial, root and total biomass and total stocked carbon during the
experiment. The reported figures are the average of the three plots per
treatment ± standard deviation. The biomass or carbon of each plot was obtained
by adding all individual pines.
La interacción clara o control × bloque tuvo un efecto
significativo en la biomasa acumulada de los pinos plantados entre 2021/22 y
2024 (F = 7.72, p < 0.001; Tabla A2 del Anexo);
no obstante, la varianza absorbida por el modelo fue pequeña (R2
ajustado = 0.089). La clara aumentó el crecimiento de
los pinos, pero su efecto dependió de la heterogeneidad ambiental (es decir,
del bloque; Tabla A2 del Anexo). El tipo de
clara (sistemática o de selección) no tuvo un efecto significativo en la
biomasa (F = 1.88, p = 0.17), siendo la variación en las claras de selección
notablemente mayor que en las claras sistemáticas (Fig. 2)
y debiéndose la variación observada sólo al efecto del bloque (F = 14.79, p
< 0.001; R2 ajustado = 0.11; Tabla A3 del
Anexo). Del mismo modo, la poda en las parcelas aclaradas no tuvo un efecto
significativo en la biomasa (F = 2.62, p = 0.11), aunque hubo una tendencia a
un mayor incremento de la biomasa en los pinos podados, y la variación
observada sólo se debió al efecto del bloque (F = 12.61, p < 0.001; R2
ajustado = 0.11; Tabla A4 del Anexo).
Vegetación leñosa espontánea
Se identificaron un total de 12 especies leñosas, excluidas
los pinos y las encinas, diferentes en los transectos de 30 m de longitud
muestreados en las 12 parcelas experimentales, además del esparto (S.
tenacissima). Esta última especie fue la dominante en las parcelas no
forestadas y el tomillo común (T. vulgaris) lo fue en las parcelas
forestadas a excepción de las parcelas con clara de selección dominadas por el
tomillo salsero (T. zygis). De manera ilustrativa, se presentan los
valores de biomasa y carbono almacenado de estas especies en el año 2021 (Tabla 3). En este año, las
parcelas no forestadas control habían almacenado dos veces más carbono en la
biomasa aérea que las parcelas forestadas control en la biomasa aérea de la vegetación
por debajo del dosel arbóreo plantado (Tabla 3).
Tabla 3. Biomasa y
carbono almacenado en los estratos de vegetación herbácea y arbustiva en los
distintos tipos de parcelas al inicio del experimento (2021).
Table 3. Biomass and stocked carbon in the herb and shrub layers of the
various experimental plot types at the start of the experiment (2021).
Señales espectrales y LiDAR
La correlación entre la altura de los pinos medida en campo
al inicio y al final del experimento y la altura medida mediante LiDAR fue muy
elevada (0.90 y 0.98 respetivamente, p < 0.001, n = 381 pinos). La
correlación entre la altura medida en campo y el NDVI fue de 0.37 y 0.17 (p
< 0.001), respetivamente, y entre el DBH medido en el campo y el NDVI fue de
0.46 (p < 0.001) y 0.16 (p = 0.0013), respetivamente (n = 381 pinos).
A nivel de parcela, la correlación entre la biomasa de los
pinos y el NDVI promedio fue baja y no significativa en ambos años (Tabla A5 del Anexo). Durante el periodo de estudio,
el NDVI promedio aumentó un 7.3 % en las parcelas forestadas control y un 14.3
% y un 2.1 %, respectivamente, en las parcelas con clara de selección y
sistemática. La correlación entre la biomasa de los pinos y el CHM promedio fue
significativa, pero la proporción de variación explicada fue baja (49 % en el
caso de la biomasa aérea; Tabla A5 del Anexo). El
CHM promedio aumentó un 8.2 % en las parcelas forestadas control y un 9.7 % y
7.9 %, respectivamente, en las parcelas con clara de selección y sistemática.
Discusión
Los objetivos de este trabajo fueron (1) identificar las
forestaciones y estimar la superficie ocupada actualmente en Campo de Montiel y
(2) evaluar cómo las forestaciones promovidas por el PFTA de la PAC con P.
halepensis y Q. ilex y las prácticas silvícolas de clara y poda en
esta zona afectan al almacenamiento de carbono. En relación con este segundo
objetivo, en general se cumplen las hipótesis de que (H1) las forestaciones
aumentan el carbono almacenado en la biomasa en relación con los matorrales
abiertos que resultan de la sucesión secundaria; (H2) las claras de las
forestaciones aumentan el crecimiento de los árboles remanentes; y (H3) la poda
de ramas secas o decrépitas aumentaron este crecimiento, aunque de forma
marginal.
Superficie forestada
Nuestro análisis indicó que la superficie con plantaciones
forestales fue pequeña en el territorio estudiado (2.7 %) y heterogénea en
cuanto al tamaño de las parcelas forestadas y especies utilizadas, dominando el
pino carrasco y la encina. La encina es la especie más abundante en el monte
mediterráneo característico de la zona y también la más plantada. Sin embargo,
el pino carrasco, inexistente de forma espontánea en la zona, ha sido la segunda
especie más utilizada para la forestación de tierras cultivadas y pastizales
debido a su tolerancia al clima del lugar y crecimiento rápido (Pausas et al., 2004b). Estas forestaciones de pinos
deben ser manejadas para crear en el medio plazo masas forestales mixtas que
sean resilientes al cambio climático, menos vulnerables a los incendios y, a su
vez, que promuevan una mayor biodiversidad (Herrero
y Zavala, 2015). Considerando la distancia a las fuentes de propágulos
naturales debido al carácter netamente agrícola de la comarca, la
transformación de estas masas monoespecíficas de pino en masas mixtas requiere
acciones de restauración activa como las plantaciones bajo cubierta combinadas
con claras que incrementen el poder facilitador del dosel del pino (Villar-Salvador, 2016; Oliet
et al., 2022; Morcillo et al., 2022). Un primer
paso sería la reposición de las marras de las encinas plantadas, que fueron
casi un 96 % en las parcelas experimentales estudiadas.
Además de la encina, otras medidas para el enriquecimiento
botánico incluirían especies productoras de frutos carnosos atractivos para los
dispersores de sus semillas (García de León et al.,
2021), como son Crataegus monogyna Jacq.,
Jasminum fruticans L., Phyllirea angustifolia L., Pyrus
bourgenana DECNE (1870), Rhamnus alataernus L., R. lyciodes L.
y Rosa spp., entre otras. Se ha observado que las claras estudiadas
en este trabajo favorecen el reclutamiento espontáneo de plántulas de encina y
de otras especies leñosas, contándose 68 individuos por ha y año en las
parcelas control y 93 y 130 individuos por ha y año en las parcelas con clara
sistemática y de selección, respectivamente (Díez-de
la Macorra et al., 2025). Además, la supervivencia temprana de las
plántulas de encina también fue favorecida por las claras (Vieco-Martínez et al., 2023).
Almacenamiento de carbono
Las parcelas forestadas almacenaron aproximadamente 30 veces
más carbono en la biomasa que las parcelas no forestadas 27 años después de la
plantación (H1). Ello se debe al lento desarrollo de la vegetación estudiada,
con fuertes limitaciones climáticas y edáficas, y al rápido crecimiento de los
pinos (Jordano et al., 2002; Lerma-Arce
et al., 2021; García de León et al., 2023).
La mayor biomasa acumulada en los pastizales no forestados en comparación con
la biomasa de los estratos de vegetación por debajo del
dosel arbóreo plantado en las parcelas forestadas no compensó la biomasa
acumulada en los pinos estudiados. No obstante, las especies herbáceas perennes
como el esparto, los caméfitos y algunos arbustos también desempeñan un papel
importante en el almacenamiento de carbono en los ambientes continentales
mediterráneos (0.44 Mg C ha-1 en las parcelas no forestadas de
nuestro estudio; Maestre et al., 2007; Montero et al., 2020). Las masas plantadas con pinos
evolucionan lentamente hacia bosques diversificados debido a la alta densidad
de estas plantaciones (en nuestro caso, ca. 541 y 441
pinos ha-1 antes y después de las claras, respectivamente) y a
la fase dinámica de exclusión en la que se encuentran (Nyland
et al., 2016). En este contexto, las claras analizadas en este estudio no
solo aumentan la disponibilidad de recursos para los árboles remanentes, sino
que también favorecen el establecimiento de un sotobosque que puede incrementar
la capacidad de almacenar carbono (Vieco-Martínez
et al., 2023).
Durante los cuatro años evaluados y en comparación con las
parcelas forestadas control, los pinos de las parcelas aclaradas tuvieron un
mayor crecimiento y almacenaron más carbono que las parcelas no aclaradas (H2),
mientras que el tipo de clara (sistemática o de selección) y la poda de las ramas bajas (H3) no afectaron al almacenamiento
de carbono. Las claras tuvieron un efecto más consistente en el crecimiento en
grosor que en altura de los árboles, confirmando los patrones característicos
generales de respuesta a la clara de árbol y rodal (Nyland
et al., 2016) que se observan también en otros casos de zonas mediterráneas
(Marchi et al., 2018; Ruiz-Peinado
et al., 2013; Navarro-Cerrillo et al., 2022;
Tabla A6 del Anexo). Las pequeñas tasas de
crecimiento pueden explicarse por el poco tiempo evaluado tras las claras, cuatro
años en las sistemáticas y tres años en las de selección.
Según este estudio, la eliminación de aproximadamente el 20
% de los pinos plantados aumentó casi un 40 % el carbono almacenado por parte
de los pinos remanentes en cuatro años (Tabla 2).
Ello sugiere una rápida compensación de la biomasa eliminada debido al efecto
de liberación del arbolado remanente que supone la clara aún en años
climáticamente más secos y calurosos de lo habitual, como
han sido los estudiados en este trabajo. Un cambio
de uso del suelo como convertir tierras agrícolas o pastizales en bosques o
plantaciones forestales sin considerar efectos indirectos puede significar la
liberación de carbono en otras regiones del planeta si se deforestan nuevos
terrenos para compensar la pérdida de producción (Powlson
et al., 2011). Este no es, sin embargo, el caso estudiado, ya que se trata
de tierras agrarias abandonadas de forma espontánea y de plantaciones poco
productivas cuyo fin primordial no es la producción de madera sino la
compensación de rentas en la población rural. En cualquier caso, en términos de
balance neto de carbono, es crítico el destino de la madera extraída de los
pinos. Este balance será más favorable, en general, si
la madera cortada se destina a usos duraderos y con alto valor de sustitución (Pukkala, 2017). Asimismo, dejar madera muerta en el
sotobosque del pinar es una opción que ralentiza la descomposición en relación
con otros usos y permite funciones de fomento de la biodiversidad como refugio
para la fauna (Bouget et al., 2012) y hábitat para
detritívoros y descomponedores (Lassauce et al., 2011).
La trituración de la madera in situ favorecerá que se descomponga más
rápidamente y el enriquecimiento orgánico del suelo (Jandl
et al., 2007). En el otro extremo, la quema como residuo o en forma de
biocombustible presentará el balance neto de carbono más desfavorable (Brack, 2017).
Capacidad predictiva de los
sensores remotos
La correlación entre la biomasa aérea de los pinos, por un
lado, y el NDVI o CHM, por el otro, que fue no significativa o significativa
con una variación absorbida relativamente baja, respectivamente, indica que
estos parámetros tienen una baja capacidad predictiva de la biomasa acumulada
en las forestaciones del área de estudio, a diferencia de lo mostrado en otros
trabajos (Calvão y Palmeirim, 2004; Galidaki et al., 2017). El pequeño número de muestras,
la heterogeneidad estructural de las forestaciones, con parches densos y huecos
y, en el caso del NDVI, las diferencias fenológicas debidas a la variación
climática pueden explicar en parte esta baja capacidad predictiva (Vázquez et al., 2013). La alta correlación entre la
altura de los pinos medida en el campo y la altura medida mediante LiDAR
sugiere la utilidad de esta última técnica para la aplicación de ecuaciones
alométricas. Sin embargo, el NDVI no resultó útil para la estimación de las
medidas biométricas. Tanto el NDVI como el CHM aumentaron entre 2022 y 2024 de
forma consistente con el aumento de la biomasa, lo que sugiere una utilidad
“cualitativa” de estos índices, más que una utilidad “cuantitativa” de
estimación de la biomasa en otras plantaciones del área de estudio. Las
investigaciones futuras deben ir dirigidas a utilizar parámetros más adecuados
obtenidos mediante teledetección y la calibración de modelos específicos para
estimar la biomasa a partir de estos.
Conclusiones e implicaciones
Nuestro trabajo permite concluir que (1) las forestaciones
del PFTA de la PAC estudiadas han aumentado ca. 30 veces el C almacenado en la
vegetación tras 27 años; (2) las claras aumentaron el carbono almacenado en el
arbolado remanente ca. un 40 % en cuatro años; (3) el
efecto significativo de las claras parece deberse sobre todo a las claras
sistemáticas, aunque la heterogeneidad ambiental y el número de réplicas no
permite conclusiones más robustas; (4) la poda de las ramas bajas aumentó
marginalmente el C almacenado en los pinos plantados; y (5) los parámetros NDVI
y CHM obtenidos mediante teledetección tuvieron una baja capacidad predictiva
de la biomasa acumulada a nivel de rodal o parcela. Estos resultados subrayan
la importancia de una gestión forestal activa para adaptarse al cambio
climático y mitigar sus efectos tal como establecen los objetivos del Plan
Nacional Integrado de Energía y Clima (PNIEC)
y del Plan Nacional de Adaptación al Cambio Climático (PNACC).
Debido a la lenta regeneración natural en la región estudiada, las
forestaciones de la PAC son capaces de almacenar más carbono por unidad de
superficie y tiempo que las expansiones naturales del bosque ligadas al
abandono rural. No obstante, como la superficie ocupada por estas plantaciones
apenas es la décima parte de la ocupada por el monte mediterráneo, el carbono
almacenado en esta formación vegetal junto al de las dehesas es previsiblemente
mayor que el almacenado en las plantaciones estudiadas.
En cualquier caso, la gestión de las forestaciones de la PAC
en clima mediterráneo continental, vulnerables al cambio climático, contribuye
a producir bosques más diversos, resilientes y con mayor capacidad de almacenar
carbono. Una cuestión por resolver es cómo y quién pagaría esta gestión de las
forestaciones para que maximicen su papel como sumideros de carbono y
proporcionen otros beneficios una vez que las ayudas del PFTA se han completado.
Dos herramientas posibles son el pago por servicios ambientales y la deducción
fiscal a los propietarios (Rey Benayas y
Bullock, 2015). Del mismo modo, la utilidad de estas forestaciones debe
considerarse en el contexto de la regeneración natural que conduce a matorrales
y bosques en buena parte de Iberia por el abandono rural. La Estrategia
de la UE sobre Biodiversidad para el año 2030 tiene el objetivo de plantar
3000 millones de árboles para abordar el cambio climático, lo que aumentaría la
cubierta forestal en alrededor de 3 millones de hectáreas. Estas plantaciones
forestales deben ser cuidadosamente diseñadas teniendo en cuenta las
características biofísicas del lugar, las especies, la densidad de plantación y
el manejo después de la plantación.
Contribución de los autores
La contribución de cada uno de los autores al trabajo es la
siguiente: Conceptualization - JMRB, JAOP; Data curation: LDM, AGP, AVM; Formal
Analysis: LDM, JMRB, AVM, LMB; Funding acquisition: JMRB, AGP; Investigation:
todos; Methodology: JMRB, AVM, JAOP; Project administration: AGP; Supervision:
JMRB; Validation: JMRB; Visualization: LDM, AVM, LMB; Writing – Original Draft:
LDM, JMRB; Writing – Review & Editing: all.
Disponibilidad de datos y código
Los datos manejados en este artículo se encuentren
accesibles en https://doi.org/10.6084/m9.figshare.30939155.
Financiación, permisos
requeridos, potenciales conflictos de interés y agradecimientos
Este estudio ha sido financiado por los proyectos “Campo de
Montiel más natural y resiliente” de la Fundación Biodiversidad, F.S.P. (Orden
TED/818/2021, convocatoria de subvenciones para la realización de proyectos que
contribuyan a implementar el Plan Nacional de Adaptación al Cambio Climático
2021-2030 en el ejercicio 2023), “REST-RES: Evaluación de técnicas de
restauración ecológica para aumentar la resiliencia frente al cambio climático
y la pérdida de biodiversidad en mosaicos forestales y agrícolas” y “ECOREST:
Evaluación de técnicas de restauración ecológica para aumentar la resiliencia
frente al cambio climático y la pérdida de biodiversidad en mosaicos
forestales, agrícolas y urbanos” del MITECO (Orden TED/898/2023 [bases
reguladoras], Orden Ministerial (extractos BOE nº 201 de 23 de agosto de 2023 y
nº 209 de 29 de agosto de 2024, respectivamente).
Las opiniones y documentación aportadas en esta publicación
son de exclusiva responsabilidad de los autores y no reflejan necesariamente
los puntos de vista de las entidades que apoyan económicamente el proyecto.
Agradecemos la ayuda de todas las personas que han
colaborado en la elaboración de material y toma de datos en este estudio:
Olivia Lorente, Clara Pemau, Carlota Arroyo, Ester Moscardó, Francisco Vieco,
Guillem Crespo, Pablo González y, especialmente, Sara Jiménez, Fernando Soneira
y Jorge García. Daniel Reinoso comentó y mejoró la redacción de los resultados
de la teledetección. Agradecemos también la colaboración de la Fundación Legado
Bustillo del Ayto. de La Solana y de Jesús Policarpo Lara Salcedo.
Los autores declaran que no tienen conflictos de interés.
Referencias
Améztegui, A., Rodrigues, M.,
& Granda, V. (2022). Uncertainty of biomass stocks in
Spanish forests: a comprehensive comparison of allometric equations. European
Journal of Forest Research, 141(3):395–407. https://doi.org/10.1007/s10342-022-01444-w.
Bouget, C., Lassauce, A., & Jonsell, M. (2012). Effects of fuelwood
harvesting on biodiversity — a review focused on the situation in Europe. Canadian
Journal of Forest Research, 42(8): 1421–1432. https://doi.org/10.1139/X2012-078.
Brack, D. (2017). Power and Heat: Impacts on the Global Climate. Chatham
House, Londres, Reino Unido. Disponible en: https://www.chathamhouse.org/2017/02/woody-biomass-power-and-heat
[consultado el 16 de abril de 2025].
Calvão, T., & Palmeirim,
J. M. (2004). Mapping Mediterranean scrub with satellite
imagery: biomass estimation and spectral behaviour. International
Journal of Remote Sensing, 25(16):3113–3126. https://doi.org/10.1080/01431160310001654978.
Castro, I., Casado, M. A.,
Ramírez-Sanz, L., De Miguel, J. M., Costa, M., & Díaz Pineda, F. (1996).
Funciones de estimación de fitomasa aérea en varias especies de matorral
mediterráneo del centro de la Península Ibérica. Orsis, 11:107–116.
Chazdon, R. L., Lindenmayer, D.,
Guariguata, M. R., Crouzeilles, R, Rey Benayas, J. M., & Lazos, E. (2020). Fostering natural forest regeneration on former agricultural land
through economic and policy interventions. Environmental Research Letters,
15:043002. https://doi.org/10.1088/1748-9326/ab79e6.
del Río, M., Pretzsch, H., Alberdi,
I., Bielak, K., Bravo, F., Brunner, A., ... Bravo-Oviedo, A. (2017).
Characterization of the structure, dynamics, and productivity of mixed-species
stands: review and perspectives. Forest Systems,
26(3):e103. https://doi.org/10.1007/s10342-015-0927-6
Díez-de la Macorra, L.,
García Pose, A., & Rey Benayas, J. M. (2025). Silvicultura restaurativa en
ecosistemas mediterráneos continentales. Fundación Internacional para la
Restauración de Ecosistemas (FIRE), Madrid, España. https://fundacionfire.org/publicacion/silvicultura-restaurativa-en-ecosistemas-mediterraneos-continentales/.
Dupont, C., Moore, B., Boasson, E. L., Gravey, V., Jordan, A., Kivimaa,
P., … von Homeyer, I. (2023). Three decades of EU climate policy: racing toward
climate neutrality? WIREs Climate Change, 15(1): e863. https://doi.org/10.1002/wcc.863.
FAO. (2025). Evaluación de los
Recursos Forestales Mundiales 2025. Food and Agriculture
Organization of the United Nations, Roma, Italia. https://www.fao.org/forest-resources-assessment/es.
Galidaki, G., Zianis, D., Gitas, I., Radoglou, K., Karathanassi, V.,
Tsakiri-Strati, M., ... Mallinis, G. (2017). Vegetation biomass
estimation with remote sensing: focus on forest and other wooded land over the
Mediterranean ecosystem. International Journal of Remote Sensing,
38(7):1940–1966. https://doi.org/10.1080/01431161.2016.1266113
García de León, D., Rey Benayas, J. M., & Andivia, E. (2021). Contributions of
hedgerows to people: a global meta-analysis. Frontiers
in Conservation Science, 2:789612. https://doi.org/10.3389/fcosc.2021.789612.
García de León, D., Rey Benayas, J. M., & Villar-Salvador, P. (2023). Assessing
the limiting factors of natural regeneration in Mediterranean planted
hedgerows. Frontiers in Ecology and Evolution, 11:1206000. https://doi.org/10.3389/fevo.2023.1206000
Frontiers.
Herrero, A., & Zavala,
M.A. (2015). Los Bosques y la Biodiversidad frente al Cambio Climático:
Impactos, Vulnerabilidad y Adaptación en España. Ministerio de Agricultura,
Alimentación y Medio Ambiente, Madrid, España. Disponible en: https://adaptecca.es/sites/default/files/editor_documentos/bosques_la_biodiversidad_frente_al_cambio_climatico_impactos_vulnerabilidad_adaptacion_2015.pdf.
Jandl, R., Lindner, M., Vesterdal, L., Bauwens, B.,
Baritz, R., Hagedorn, F., ... Byrne, K. A. (2007). How strongly can forest
management influence soil carbon sequestration? Geoderma,
137(3–4):253–268. https://doi.org/10.1016/j.geoderma.2006.09.003.
Jordano, P., García, C., Godoy, J.
A., & García-Castaño, J. L. (2002). Reclutamiento de especies leñosas en
bosques tropicales: la importancia de la dispersión de semillas. Ecosistemas,
11(1):45–60. https://www.revistaecosistemas.net/index.php/ecosistemas/article/view/308.
Lang, N., Kalischek, N., Armston, J., Schindler, K., Dubayah, R.,
Wegner, J.D. (2022). Global canopy height regression and uncertainty estimation
from GEDI LIDAR waveforms with deep ensembles. Remote Sensing of Environment,
268:112760.
Lassauce, A., Paillet, Y., Jactel, H., & Bouget, C. (2011). Deadwood as
a surrogate for forest biodiversity: Meta-analysis of correlations between
deadwood volume and species richness of saproxylic organisms. Ecological
Indicators, 11(5):1027–1039. https://doi.org/10.1016/j.ecolind.2011.02.004.
Legesse, F., Degefa, S., & Soromessa, T. (2024). Estimating carbon
stock using vegetation indices and empirical data in the upper awash river
basin. Discover Environment 2:137. https://doi.org/10.1007/s44274-024-00165-8.
Lerma-Arce, V.,
Oliver-Villanueva, J. V., Segura-Orenga, G., & Urchueguia-Schölzel, J. F.
(2021). Comparison of alternative harvesting systems for
selective thinning in a Mediterranean pine afforestation (Pinus halepensis
Mill.) for bioenergy use. iForest, 14(5):465. https://doi.org/10.3832/ifor3636-014.
Leyva Cabello, F.,
& Matas González, J. (2013). Mapa Geológico de España -- Hoja nº
787. 1:50.000. Segunda Serie (MAGNA), Primera Edición. Instituto Geológico
y Minero de España (IGME), Madrid, España.
Locatelli, B., Pavageau, C., Pramova, E., & Di
Gregorio, M. (2015). Integrating climate change mitigation and adaptation in
agriculture and forestry: Opportunities and trade-offs. Wiley Interdisciplinary Reviews: Climate Change, 6(6):585–598. https://doi.org/10.1002/wcc.357.
Maestre, F. T., Ramírez, D. A.,
& Cortina, J. (2007). Ecología del esparto (Stipa tenacissima L.) y
los espartales de la Península Ibérica. Ecosistemas 16(2). https://www.revistaecosistemas.net/index.php/ecosistemas/article/view/89.
Marchi, E., Chung, W., Visser, R., Abbas, D., Nordfjell, T., Mederski, P. S.,
... Laschi, A. (2018). Sustainable Forest Operations (SFO): a new paradigm in a
changing world and climate. Science of the Total Environment,
634:1385–1397. https://doi.org/10.1016/j.scitotenv.2018.04.084.
Menéndez-Miguélez, M.,
Calama, R., del Río, M., Madrigal, G., López-Senespleda, E., Pardos, M., &
Ruiz-Peinado, R. (2022). Species-specific and generalized
biomass models for estimating carbon stocks of young reforestations. Biomass
and Bioenergy, 161:106453. https://doi.org/10.1016/j.biombioe.2022.106453.
Ministerio de Agricultura, Pesca y
Alimentación (1993). Real Decreto 378/1993, de 12 de marzo, por el que
se establece un régimen de ayudas para fomentar inversiones forestales en
explotaciones agrarias y acciones de desarrollo y aprovechamiento de los
bosques en las zonas rurales. BOE núm. 76, de 30 de marzo de 1993,
páginas 9378 a 9385. https://www.boe.es/eli/es/rd/1993/03/12/378.
Montero, G., López-Leiva, C.,
Ruiz-Peinado, R., López-Senespleda, E., Onrubia, R., & Pasalodos, M.
(2020). Producción de biomasa y fijación de carbono por los matorrales
españoles y por el horizonte orgánico superficial de los suelos forestales.
Ministerio de Agricultura y Alimentación-INIA, Madrid, España.
Morcillo, L., Turrión, D.,
Soliveres, S., Chirino, E., Vallejo, R. V., & Vilagrosa, A. (2022). Moderate pine cover maximizes 10-year survival and growth in
late-successional species of contrasting functional strategies. Forest
Ecology and Management, 509:120098. https://doi.org/10.1016/j.foreco.2022.120098.
Moreau, G., Auty, D., Pothier, D., Shi, J., Lu, J., Achim, A., & Xiang,
W. (2020). Long-term tree and stand growth dynamics after thinning of various
intensities in a temperate mixed forest. Forest Ecology and
Management, 473:118311. https://doi.org/10.1016/j.foreco.2020.118311.
Navarro-Cerrillo, R. M.,
Ruiz-Gómez, F. J., Camarero, J. J., Castillo, V., Barberá, G. G., Palacios- Rodríguez, G., ... del Campo,
A. D. (2022). Long-term carbon sequestration in pine forests
under different silvicultural and climatic regimes in Spain. Forests,
13(3):450. https://doi.org/10.3390/f13030450.
Nyland, R. D., Kenefic, L. S., Bohn, K. K., & Stout, S.L. (2016). Sylviculture.
Concepts and applications. Waveland Press Income, Illinois, USA.
Oliet Palá, J. A., Bravo Fernández, A., Rubio Sánchez, A., de Frutos López,
S., & Roig Gómez, S. (2022). Descripción del dispositivo
experimental del proyecto FORADMIT: acciones selvícolas para la diversificación
de pinares artificiales. Fundación Conde del Valle de Salazar, Madrid,
España. https://oa.upm.es/80752/.
Pausas, J. G., Blade, C., Valdecantos, A., Seva, J. P., Fuentes,
D., Alloza, J. A., … Vallejo, R.
(2004a). Pines and oaks in the restoration of Mediterranean landscapes of
Spain: New perspectives for an old practice – a review. Plant Ecology,
171:209–220. https://doi.org/10.1023/B:VEGE.0000029381.63336.20.
Pausas, J. G., Bradstock, R. A., Keith, D. A., & Keeley, J.E. (2004b).
Plant functional traits in relation to fire in crown-fire ecosystems. Ecology,
85(4):1085–1100. https://doi.org/10.1890/02-4094.
Posit
Team. (2024). RStudio:
Integrated development environment for R (versión 2024.12.1.563)
[Software]. Posit Software, PBC. https://www.posit.co/.
Powlson, D.S., Whitmore, A. P., & Goulding, K. W. T. (2011). Soil
carbon sequestration to mitigate climate change: a critical re-examination to
identify the true and the false. European Journal of Soil Science,
62:42–55.
Pukkala, T. (2017). Does management improve the carbon
balance of forestry? Forestry, 90(1):125–135.https://www.cabidigitallibrary.org/doi/full/10.5555/20173021319.
R
Core Team. (2023). R:
A language and environment for statistical computing. R Foundation for
Statistical Computing, Vienna, Austria. https://www.R-project.org/.
Rey
Benayas, J. M., & Bullock, J. M. (2015).
Vegetation restoration and other actions to enhance wildlife in European
agricultural landscapes. En: Pereira, H.M., Navarro, L.M. (eds.), Rewilding
European Landscapes, pp. 127-142. Springer International Publishing, Cham, Switzerland.
https://doi.org/10.1007/978-3-319-12039-3_7.
Rivero, R.G., Fernández, R.C.,
& Montes, R.I. (2016). Defensa y prevención de incendios forestales.
Síntesis, Madrid, España. ISBN Papel: 9788490773062 / ISBN eBook:
9788490778227. Disponible en: https://www.sintesis.com/libro/defensa-y-prevencion-de-incendios-forestales.
Ruiz-Peinado, R., del Río, M.,
& Montero, G. (2011). New models for estimating the carbon
sink capacity of Spanish softwood species. Forest Systems,
20(1):176–188. https://doi.org/10.5424/fs/2011201-11643.
Ruiz-Peinado, R.,
Bravo-Oviedo, A., López-Senespleda, E., Montero, G., & del Río, M. (2013). Do thinnings influence biomass and soil carbon stocks in
Mediterranean maritime pinewoods? European Journal of Forest Research,
132(2):253–262. https://doi.org/10.1007/s10342-012-0672-z.
Simard, M., Pinto, N., Fisher, J. B., & Baccini, A. (2011). Mapping
forest canopy height globally with spaceborne lidar. Journal of Geophysical
Research 116:G04021. https://doi:10.1029/2011JG001708.
Suganuma, H. S., Kawada, K., Smaout, A., Suzuki, K., Isoda, H., Kojima, T.,
& Abe, Y. (2012). Allometric equations and biomass
amount of representative Tunisian arid land shrubs for estimating baseline. Journal
of Arid Land Studies, 22(1):219–222. https://doi.org/10.1186/s40663-015-0053-4.
Vadell, E., de Miguel, S., Centeno, G. F., Robla, E., Cuzzi, M. L., &
García, J. P. (2019). La forestación de tierras agrícolas: balance de un
instrumento de política forestal para el cambio del uso de la tierra. Cuadernos
de la Sociedad Española de Ciencias Forestales, 45:1–20. https://doi.org/10.31167/csecfv0i45.19497.
Vayreda, J., Martínez-Vilalta, J.,
Gracia, M., & Retana, J. (2012). Recent climate changes
interact with stand structure and management to determine changes in tree
carbon stocks in Spanish forests. Global Change Biology,
18(3):1028–1041. https://doi.org/10.1111/j.1365-2486.2011.02606.x.
Vázquez, P., Adema, E., &
Fernández, B. (2013). Dinámica de la fenología de la vegetación a partir de
series temporales de NDVI de largo plazo en la provincia de La Pampa. Ecología
Austral, 23(2):77‑86.
https://doi.org/10.25260/EA.13.23.2.0.1163.
Vicente-Serrano, S. M.,
López-Moreno, J. I., Beguería, S., Lorenzo-Lacruz, J., Sánchez-Lorenzo, A.,
García-Ruiz, ... Espejo, F. (2014). Evidence of increasing
drought severity caused by temperature rise in southern Europe. Environmental
Research Letters, 9(4):044001. https://doi.org/10.1088/1748-9326/9/4/044001.
Villar-Salvador, P. (2016). Restoration of Spanish pine plantations: A main
challenge for the 21st century. Reforesta, 1(1):53-66. https://doi.org/10.21750/REFOR.1.04.4.
Vieco-Martínez, A.,
Rey-Benayas, J. M., Oliet, J. A., Villar-Salvador, P., & Martínez-Baroja,
L. (2023). Efectos de la forestación de tierras agrícolas mediterráneas y de su
manejo en el establecimiento de árboles y arbustos. Ecosistemas, 32(especial):2460. https://doi.org/10.7818/ECOS.2460.
Anexo / Appendix
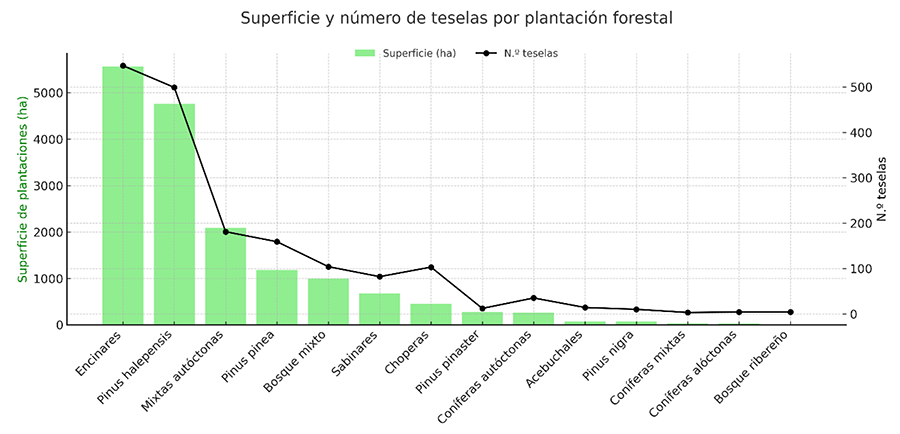
Figura A1. Número de teselas y superficie (ha)
de cada tipo de plantación forestal en Campo de Montiel.
Figure A1. Number of and area (ha) of every tree plantation type in Campo de
Montiel.
Tabla A1. Valores
promedio (± desviación estándar) de la altura (m) y el DBH (cm) en las parcelas
forestadas; n = 366.
Table A1. Mean (± standard deviation) of height (m) and DBH (cm) in the
afforested plots; n = 366.
Tabla A2. Comparaciones
múltiples de Tukey para la interacción tratamiento (claras o control) × bloque
en la diferencia de biomasa. Para el modelo completo, F5,360
= 8.11, p <0.001.
Table A2. Tukey's multiple comparisons for the treatment (thinning or
control) × block interaction on biomass difference. For the full model, F5,360
= 8.11, p < 0.001.
Nota: Valores positivos indican que la diferencia
estimada de la biomasa aumentó más en la primera interacción que en la segunda.
Valores negativos indican que la biomasa aumentó más en la segunda interacción
que en la primera. En negrita se destacan las comparaciones con
diferencias significativas. Las diferencias estimadas están en la escala de 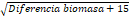 .
.
Fórmula del modelo: lm( ~
tratamiento (claras o control) × bloque)
Tabla A3. Comparaciones
múltiples de Tukey para la interacción tipo de clara (selección o sistemática)
× bloque en la diferencia de biomasa. Para el modelo completo,
F3,322 = 10.38, p <0.001.
Table A3. Tukey's multiple comparisons for the thinning type (selection or
systematic) × block interaction on biomass difference. For the full model, F3,322
= 10.38, p < 0.001.
Nota: Valores negativos indican que la diferencia
estimada de biomasa aumentó más en el primer término de la comparación que en
el segundo. En negrita se destacan las comparaciones con diferencias
significativas.
Fórmula del modelo: lm(Diferencia biomasa ~ tipo de
clara (sistemática o selección) + bloque).
Tabla A4. Comparaciones
múltiples de Tukey para la interacción poda (sí o no) × bloque en la diferencia
de biomasa. Para el modelo completo, F3,322 =
10.66, p <0.001.
Table A4. Tukey's multiple comparisons for the pruning (yes or no) × block
interaction on biomass difference. For the full model, F3,322
= 10.66, p < 0.001.
Nota: Valores negativos indican que la diferencia
estimada de biomasa aumentó más en el primer término de la comparación que en
el segundo. En negrita se destacan las comparaciones con diferencias
significativas.
Fórmula del modelo: lm(Diferencia biomasa ~ poda
(sí o no) + bloque).
Tabla A5. Matriz
de correlación de Pearson de la biomasa total y la aérea de los pinos, el
parámetro CHM obtenido a partir del LiDAR y el NDVI obtenido a partir de las
señales multiespectrales en las parcelas reforestadas estudiadas.
Table A5. Pearson correlation matrix of the total and aboveground biomass of
pines, the CHM parameter obtained from LiDAR and the NDVI obtained from the
multispectral signals in the afforested plots studied.
Tabla A6. Cifras de
biomasa o secuestro de carbono reportadas por otros estudios de la literatura
científica. Para la búsqueda sistemática, se dieron las siguientes
instrucciones a ChatGPT: (1) “Necesito artículos científicos con las siguientes
palabras clave: biomass, carbon sequestration, Mediterranean, plantation,
tree”; (2) “completa la información de la tabla adjunta”; (3) para cada
artículo individual, “resume el artículo resaltando la información de la tabla”
(esta segunda revisión amplió la información de la instrucción (2)).
Complementariamente, los artículos identificados proporcionaron otros estudios
con cifras de interés.
Table A6. Biomass or carbon sequestration figures reported by other studies
in scientific literature. For the systematic search, ChatGPT was given the
following instructions: (1) “I need scientific articles with the keywords:
biomass, carbon sequestration, Mediterranean, plantation, tree”; (2) “Complete
the information in the attached table”; (3) For each individual article,
“Summarize the article and highlight the information in the table” (this second
review expanded on the information in instruction (2)). In addition, the
identified articles provided other studies with figures of interest.
|
Referencia
|
Tipo de sistema
forestal
|
Tipos de tratamiento
|
Biomasa/secuestro de
C
|
|
Ruiz-Peinado
et al. 2017
|
Plantaciones de Pinus y Quercus
|
Diferentes edades y especies para distintos tipos de
tratamientos silvícolas (claras, periodos de rotación, etc.).
|
Sin clara: 145 Mg C/ha
Clara ligera: 151 Mg C/ha
Clara fuerte: 116 Mg C/ha
|
|
Ruiz-Peinado et al. 2013
|
Plantaciones de Pinus pinaster
|
Plantaciones de 59 años.
Claras de distintas intensidades.
|
Sin clara: 196.9 Mg C/ha
Clara moderada: 139.3 Mg C/ha
Clara intensa: 126.8 Mg C/ha
|
|
Ruiz-Peinado et al. 2016
|
Plantaciones de Pinus sylvestris L.)
|
Plantaciones de 52 años.
Claras con rotaciones de 10 años.
|
Sin clara: 206.8 Mg C/ha
Clara moderada: 164 Mg C/ha
Clara intensa: 148.8 Mg C/ha
|
|
Bravo-Oviedo
et al. 2015
|
Pinus sylvestris L.
(naturales)
|
Sistemas aclarados (baja intensidad y moderada) vs. sin
aclarar.
Rodales de 90 años.
|
Sin clara: 129.2 Mg C/ha
Clara intensidad baja: 105.9 Mg C/ha
Clara intensidad moderada: 93.3 Mg C/ha
|
|
Navarro-Cerrillo
et al. 2022
|
Plantaciones y masas naturales maduras de pino (nueve Pinus spp.)
|
Claras de intensidad baja y alta
|
P. halepensis
Sin clara: 94.5 Mg C/ha
Clara baja intensidad: 64.1 Mg C/ha
Clara alta intensidad: 49.9 Mg C/ha
|
|
del Río et al.
2017
|
Bosques de pinos mediterráneos (P. sylvestris, P. pinaster,
P. pinea)
|
Rodales coetáneos vs multietáneos con aclareos durante 100
años
|
Coetáneo: 145 Mg C/ha en 100 años;
Multietáneo: 130 Mg C/ha
Multietáneo fija más C anual (1.334 vs. 1.195 Mg C/ha/año)
|
|
Erkan et al. 2023
|
Plantacion de Pinus brutia Ten.
|
Claras moderadas, intensas y sin aclareos
|
Sin clara: 151.1 Mg C/ha
Clara moderada: 132.5 Mg C/ha
Clara intensa: 133.3 Mg C/ha
|
|
Cachinero-Vivar
et al. 2021
|
Plantaciones de pinos mediterráneos (40-42 años)
|
Parcelas sin claras, con claras moderadas y claras intensas
|
P. pinaster
Sin clara: 33.2 Mg C/ha
Clara moderada: 49.6 Mg C/ha
Clara intensa: 73.1 Mg C/ha
P. nigra
Sin clara: 29.0 Mg C/ha
Clara moderada: 33.0 Mg C/ha
Clara intensa: 36.4 Mg C/ha
P.sylvestris
Sin clara: 38.5 Mg C/ha
Clara moderada: 43 Mg C/ha
Clara intensa: 51.5 Mg C/ha
|
Referencias del Anexo
Bravo-Oviedo, A.,
Ruiz-Peinado, R., Modrego, P., Alonso, R., & Montero, G. (2015). Forest thinning impact on carbon stock and soil condition in
Southern European populations of P. sylvestris L. Forest Ecology and
Management 357:259–267. https://doi.org/10.1016/j.foreco.2015.08.005
Cachinero-Vivar, A. M.,
Palacios-Rodríguez, G., Lara-Gómez, M. A., & Navarro-Cerrillo, R. M. (2021).
Thinning effect of C sequestration along an elevation gradient
of Mediterranean Pinus spp. plantations. Forests
12(11):1583. https://doi.org/10.3390/f12111583
del Río, M., Barbeito, I.,
Bravo-Oviedo, A., Calama, R., Cañellas, I., Herrero, C., … Bravo,
F. (2017). Mediterranean pine forests: management effects on carbon stocks. En:
Bravo, F., LeMay, V., Jandl, R. (eds.), Managing Forest Ecosystems: the
challenge of climate change, pp. 301–327. Springer, Cham, Suiza. https://doi.org/10.1007/978-3-319-28250- 3_15
Erkan, N., Güner, Ş. T., & Aydın, A. C. (2023). Thinning effects on
stand growth, carbon stocks, and soil properties in Brutia pine
plantations. Carbon Balance and Management 18(1):6. https://doi.org/10.1186/s13021-023-00226-0
Navarro-Cerrillo, R. M.,
Ruiz-Gómez, F. J., Camarero, J. J., Castillo, V., Barberá, G. G., Palacios-
Rodríguez, G., ... del Campo, A. (2022). Long-term carbon
sequestration in pine forests under different silvicultural and climatic
regimes in Spain. Forests 13(3):450. https://doi.org/10.3390/f13030450
Ruiz-Peinado, R.,
Bravo-Oviedo, A., López-Senespleda, E., Montero, G., & del Río, M. (2013). Do thinnings influence biomass and soil carbon stocks in
Mediterranean maritime pinewoods? European Journal of Forest Research
132(2):253–262. https://doi.org/10.1007/s10342-012-0672-z
Ruiz-Peinado, R., Bravo-Oviedo, A., Montero, G., & del Río, M. (2016). Carbon
stocks in a Scots pine afforestation under different thinning intensities
management. Mitigation and Adaptation Strategies for Global Change
21:1059–1072. https://doi.org/10.1007/s11027-014-9585-0
Ruiz-Peinado, R.,
Bravo-Oviedo, A., López-Senespleda, E., & del Río, M. (2017). Forest management and carbon sequestration in the Mediterranean
region: a review. Forest Systems 26(2):eR04S. https://doi.org/10.5424/fs/2017262-11205