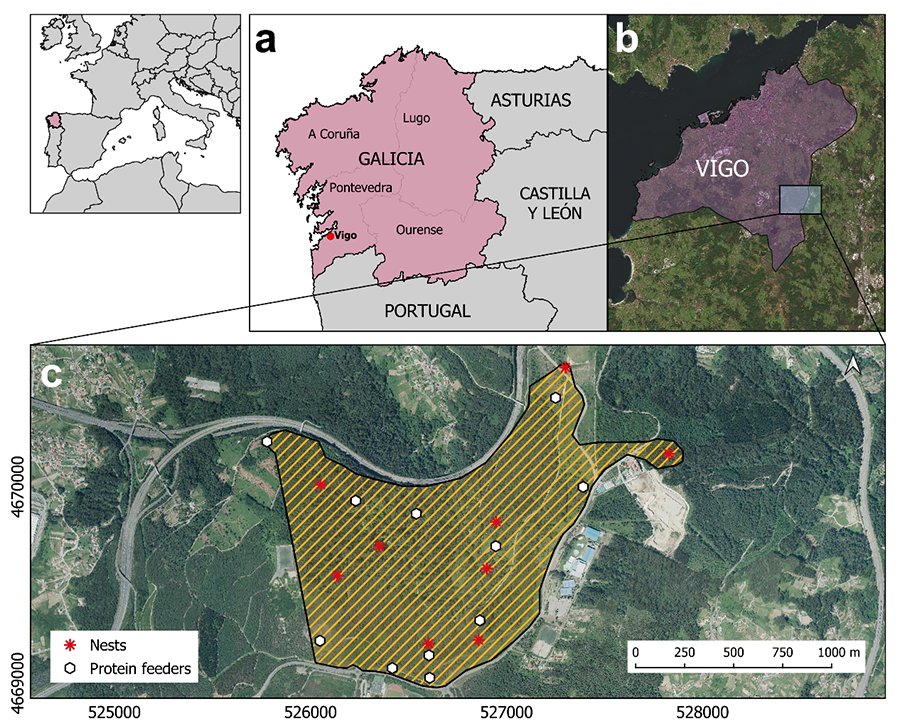
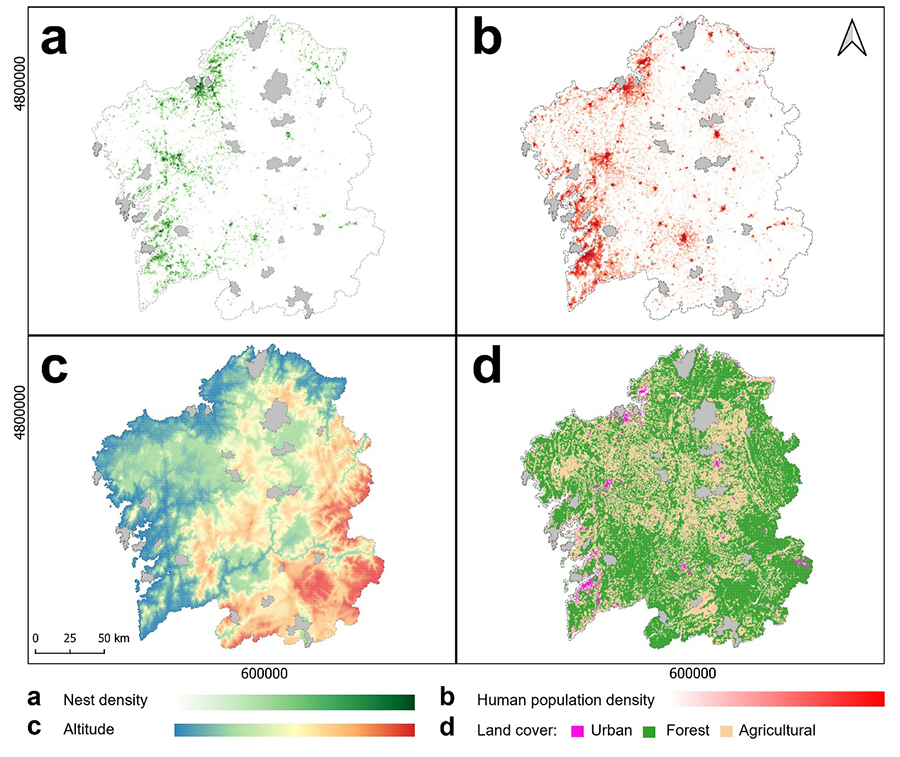
|
GAM model
|
|
Parametric
coefficients
|
Estimate
|
Std. Error
|
z value
|
Pr(>|z|)
|
|
Intercept
|
3.32
|
0.032
|
103.13
|
< 0.001 ***
|
|
Nest
2
|
-1.25
|
0.160
|
-7.82
|
< 0.001 ***
|
|
Nest
3
|
0.71
|
0.041
|
17.39
|
< 0.001 ***
|
|
Smooth
terms
|
edf
|
Ref. df
|
Chi. sq
|
p-value
|
|
S(Date):
Nest 1
|
3.86
|
4.59
|
300.8
|
< 0.001 ***
|
|
S(Date):
Nest 2
|
5.79
|
6.53
|
840.4
|
< 0.001 ***
|
|
S(Date):
Nest 3
|
1.02
|
1.04
|
664.4
|
< 0.001 ***
|
Nest 3 had the highest activity throughout the studied season, with the
highest overall activity on the first day of recording (Fig. 5). Its activity decreased by 87 %
between the first and last day. Nest 2 was the second most active nest during
the first set of recordings but then showed a sharp (70 %) decline in activity,
which stopped completely during the final set of recordings. Nest 1 had the
least variation in activity over time. Its maximum activity was achieved during
the second set of recordings, and at the end of the recordings it reduced its
activity by 86 %.
The coefficient of variation increased as
nest activity decreased with a recording time of 5 minutes (Fig 5). The coefficient of variation
for nest 2 is not shown for the final recording set as only a single exit was
registered in any 5-minute video in that set.
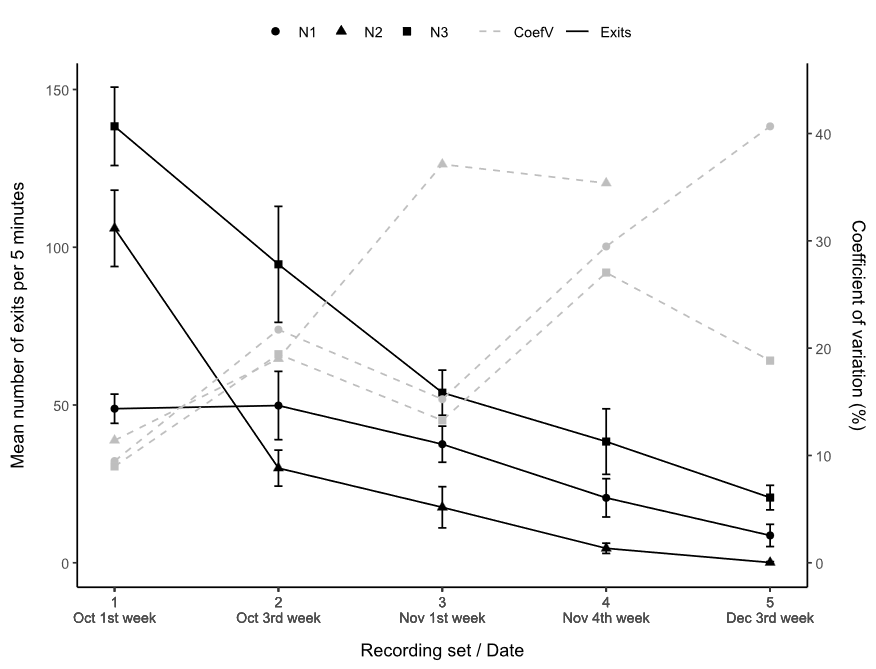
Figure 5.
Relation between nests exits (Exits) (black continuous line) and coefficient of
variation (CoefV) (grey dotted line) of the three nests studied N1 (●), N2 (▲),
and N3 (■) grouped by recording sets. Error bars represent standard
deviation of data (N=12).
Figura 5. Relación
entre las salidas del nido y el coeficiente de variación (CoefV) de los tres
nidos estudiados (N1, N2 y N3) agrupados por grupos de grabación. Las barras de
error representan la desviación estándar de los datos (N=12).
Discussion
Factors related to citizen
reported nest data distribution
Human population density and urban land
cover are the variables that best explain the likelihood of finding nests in
Galicia. The most probable cause for this fact lies in the method of nest
registration itself. The Xunta de Galicia's database consists of nest locations
reported by citizens. In non-urban areas, the human population density is lower
and consequently the number of observers, decreasing the likelihood of nests
being discovered (Franklin
et al. 2017). Moreover, nests in forested
areas are more difficult to see as they are found in the treetops and
camouflaged among the foliage (Rodríguez-Flores et al. 2019).
Several studies (Villemant
et al. 2011b; Choi et al. 2012b; Rodríguez-Flores et al. 2019) also suggest a greater preference by the hornet for urban
environments. Thus, the high densities of nests found in urban areas are likely
linked to both the number of observers and to the hornet's habitat preferences.
Moreover, agricultural land cover favours nest observations, but its influence
is almost four times lower than that of urban areas
(Supplementary Information Tabla A1)
probably due to the lower human population density and
the nesting placement. Nests in cities receive more attention from the public,
even the smallest ones are reported, whereas reports of harmless nests in rural
areas are less likely to occur (Choi
et al. 2019). In addition, city dwellers
encounter nests closer to their homes, leading to an increased perception of
risk, also fostered by the media (Do
et al. 2019). In this context, Pazos et al. (2022) found that although people in Galicia report notifying authorities
about the presence of nests for management purposes, the removal services
report that most of the calls are justified by alleged allergies to the species
or health concerns. This suggests a higher demand for the removal of nearby
nests driven by fear of the species.
Altitude is also a good indicator of the
likelihood of a nest being discovered, with higher altitudes decreasing the
odds of nest encounters. This may be explained by the species expanding rapidly
through coastal areas and preferring to nest in mild climates at low altitudes (Rodríguez-Flores et al. 2019). Nests are often found under 800 meters above sea level (Archer 2006)
despite that they can be present at altitudes as high as 1100 meters above sea
level (Porporato et al.
2014). In turn, human populations in Galicia are
concentrated in coastal areas or associated with large cities and transport
routes (IGE 2022). Therefore, increasing altitude decreases both the probability
that a nest is present and reported by an observer.
The large number of nest reports indicates
the wide distribution of the species in Galicia and the commitment of the
community. The latter is crucial to help administrations manage the invasion
effectively; however, many people are still misinformed, in part due to
sensationalism or fake news (Pazos
et al. 2022). The implementation of public
information programmes on the management of the species would improve
cooperation between citizens and authorities and optimise the resources
allocated to the species control, saving costs and time.
Comparison of reported nest
densities with those found by means of radio-tracking in a coniferous forest
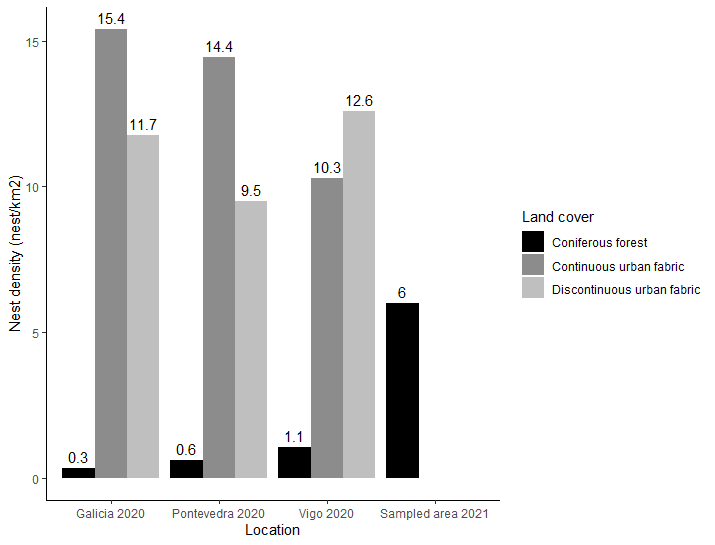
According to the Xunta de Galicia database,
nest density in the municipality of Vigo in 2020 was 10 to 12 times higher in
the urban environment (~11 nests/ km2) compared to coniferous forest
(~1 nest/ km2). In France, Villemant et al. (2011b) reported
that 49 % of the nests found were in the urban environment, while only 7 % in
forest, resulting in 7-fold higher density in urban than forest environments.
In the community of Andernos-les-Bains (France) almost all reported nests were
found in the urban area with a density of 10 nests/km2 (Franklin et al. 2017). Closer to Galicia, in Portugal, Carvalho et al. (2020) found a
lower nest density in the urban environment of 5.4 ±3.3 nests/km2.
The results obtained in this study from an
exhaustive radio-tracking search conducted in the summer of 2021 on outskirts
of Vigo showed a nest density in the coniferous forest of 6 nests/km2.
This is more than five times the density reported in the official data for the
municipality of Vigo in coniferous forests. However, the sampling probably did
not detect all the nests in the radio-tracked area. Using the method based on
directions and timings (Rojas-Nossa
et al. 2022) we can estimate that the actual
number of nests was greater than that determined by radio-tracking in the
summer of 2021. Thus, it can be concluded that the Xunta de Galicia database
may be greatly underestimating nest density in coniferous forests. This
pattern can probably be explained by the scarcity of observers in forest areas
and the fact that perennial coniferous forests increase the likelihood of not
finding a nest even in autumn compared to deciduous forests (Bessa et al. 2016). These hypotheses are supported by the fact that no nests were
reported the previous year in the sampled area.
The reported density of nests in the
coniferous forest was higher in Vigo than in the province of Pontevedra, which
in turn was higher than that for Galicia overall. This can be explained by
reported nest density being influenced by human population density. The human
population density of Vigo is higher than that of Pontevedra, which in turn is
higher than that for Galicia overall. In addition, the nesting preference for
coastal areas, the continental climate of inland Galicia, topography (especially
mountains) or the different stages of the invasion may also contribute to this
distribution pattern (Robinet
et al. 2017; Rodríguez-Flores
et al. 2019; Carvalho et al. 2020).
The high abundance of nests in forested
areas makes us hypothesise that it could act as a source for the species. In
this way, the higher number of queens caught in forests during spring found by Rojas-Nossa et al. (2018) supports this hypothesis. Management measures generally focus on
urban environments, trying to reduce hornet densities, but nearby forests can
contribute to increasing it in urban centres. Although the dispersion ability
of queens is poorly documented, Beggs
et al. (2011) reported that queens can fly
about 30 km in 1 day, making the colonisation of urban areas and nest
installation from nearby forests plausible. In addition, V. velutina
workers may appear in cities because they can forage at distances of up to 1 km
from the nest (Poidatz
et al. 2018; personal observation AL). However,
the management of the invasion in these areas is not easy due to the absence of
observers and the fact that nests are hidden in the trees (Bessa et al. 2016). In this way, the management of forests near cities must be considered
by the administrations to reduce the impact of the invasion in the cities.
In addition, Vigo is the largest city in
Galicia, containing extensive areas of continuous urban fabric far from green
areas. Nest density in the municipality is greater in discontinuous urban areas
than in continuous ones. The presence of green zones interspersed in the urban
environment could favour the establishment of the species. These areas contain
higher biodiversity, and hornets have a greater number of potential food
sources and nesting sites, which can be limiting in the continuous urban fabric
(Choi et al. 2012a).
Monitoring nests activities
The
three nests monitored in the study reduces their activities during the late
season; however different behaviours were observed. Both nests 2 and 3
demonstrated a sudden drop in activity between the first two recording sets
being more accused in nest 2. This may be because it was located at a height of
45 metres and quite far from the tree trunk. These characteristics suggest that
weather factors (mainly rain and wind) could exert greater pressure on the nest
because it was unprotected (Matsuura and Yamane 1990). Such a location also favours
greater exposure to the sun, which can increase the nest temperature and
negatively influence the colony (Rodríguez-Flores et al. 2019). The decrease in activity was persistent
as it was the only nest that collapsed during the recording period showing
clear signs of deterioration (changes in colour and damage to the outer nest
structure; personal observations AL).
Otherwise,
the population of V. velutina nests is closely related to the size of
the nest (Rome
et al. 2015). This may explain the higher activity of nest 3, as it was
visually the largest of the three nests and the lower activity observed in nest
1. The abundant vegetation and the location close to the tree trunk protect V.
velutina nests from the increasingly intense environmental factors as the
season progressed too.
The
proximity of the three nests to each other (at a maximum distance of 470 meters
from one another) excludes the possibility that the differences observed in
activity are due to climate or altitude. The results confirm the influence of
the recording date on the activity of the nests, which collapsed later in
January. This is in line with the life cycle of the species, in which all
individuals (except hibernating gynes) die with the onset of winter (Monceau et al.
2014).
Activity results are similar to those found in previous work in southwestern
France (Rome et al.
2015; Monceau et al. 2017). The maximum levels of activity in these
studies occurred in mid/late October, coinciding with the activity shown by nest
1. In nests 2 and 3 this peak of activity occurs at the beginning of the
recording period, in late September and early October or even earlier. The
higher variability observed in nest activities make us consider that the
location of the nest is crucial for the evolution of hornet’s activity.
Protected nests seem to be bigger and more active than unprotected ones. Also
important, is the location of embryo nests as they are more vulnerable to the
climatic factors (Diéguez-Antón
et al. 2022) and may influence
the future development of the secondary nest.
Nest activities were confirmed in late autumn and even in early winter. Related
to this, Feás
Sánchez and Charles (2019) support that it is
extremely important to remove V. velutina nests during the winter as
well as the rest of the year.
To
estimate nest activity, the observation time of 5 minutes proposed by Monceau et al. (2017) seems adequate up to the first
recording set in this study. However, with reduced nest activity, the
coefficient of variation between recordings increases to around 30 % by the
final recordings, exceeding it in some cases, such that the confidence level
with 5-minute recordings becomes insufficient (Gordón-Mendoza and
Camargo-Buitrago 2015). Consequently, it is desirable to increase the recording
time for activity at nests late in the season to obtain a higher level of
confidence (coefficient of variation < 30 %). It is important to note that
in addition to recording time, the day and time of sampling should be
considered when comparing activities, as these factors also determine nest
activity (Williams 1988). In this study,
recordings were made only on sunny days and at midday to observe the peak daily
activity. Consequently, understanding the temporal dynamics of nest collapse
allows extending nest removal operations further into the late season,
potentially reducing the spread of the invasion. However, since not all nests
follow the same activity patterns, the monitoring of nest exits for a duration
exceeding five minutes under favourable weather conditions can help identify
inactive nests and avoid unnecessary removal efforts and associated costs.
These
data are a first effort to establish decision criteria for nest removal at the
end of the species' annual cycle. Although the sample size in our study is
limited, the observed trends are similar to those reported in southern France (Monceau et al.
2017).
Improving decision capacities contribute to reduce the economic costs of the
invasion management. In this way, the total budget to manage the invasion
overcame € 2 million in the community of Galicia in the year 2020 (Xunta de Galicia 2020a) and beekeepers receive
government aid to compensate for hornet losses up to € 700 000 more (Xunta de Galicia 2020b). Moreover, detrimental
effects on pollination services and behaviour of pollinators were demonstrated
by Rojas-Nossa
et al. (2022). In addition, the invasion entails human health
costs due to hornet stings. All this data supports the urgency of acting
against the invasion as it presented serious economic, ecosystemic and human
health problems.
Conclusions
The official databases concerning V.
velutina nests in Galicia are compiled using citizen reports. In this way,
they do not offer a representative information of the distribution and
abundance of the hornet in all the Galician territory. This study showed that citizen-reported
data are mainly biased by the presence of observers and urban land use but may
be related to other variables like altitude. In this way, nests located in
urban areas are well represented in citizen reports which indicates that community
engagement is effective in locating nests. However, nests in forestry land are
underrepresented due to the near absence of observers and how hidden the nests
are. Forests can act as a source for the species in the invaded area.
Consequently, at the invasion front, greater search efforts in forest must be
considered by the authorities to reduce the advance of the species.
To accurately assess nest activity, it is
recommended to enhance the duration of time recording when a decline in
activity is observed. Observations made in this study support extending efforts
in nest removal into late autumn. This information can be useful to optimize
resources for managing the V. velutina invasion and reduce socioeconomic
and ecosystem impacts.
Author´s contribution
Aarón Lagoa: Conceptualization; Data curation; Formal Analysis; Investigation;
Methodology; Visualization; Writing – Original Draft; Writing – Review &
Editing. Iria Villar: Conceptualization; Data curation; Formal Analysis;
Investigation; Methodology; Visualization; Writing – Original Draft; Writing –
Review & Editing. Sandra V. Rojas-Nossa: Conceptualization; Formal
Analysis; Investigation; Methodology; Visualization; Writing – Review &
Editing. Peter J. Kennedy: Conceptualization; Data Curation; Investigation;
Methodology; Visualization; Writing – Review & Editing. Salustiano Mato:
Conceptualization; Supervision, Writing – Review & Editing. Josefina
Garrido: Conceptualization; Funding
Acquisition; Supervision; Writing – Review & Editing.
Data availability
The datasets
used during the current study were provided on demand to the Regional
Government of Galicia. More information could be required on reasonable
request.
Financing, required permits, potential
conflicts of interest and acknowledgments
This work was financed by the Interreg
Atlantic Area Program (European Regional Development Fund—ERDF, European
Union): EAPA_800/2018—Atlantic-Positive. The authors declare no conflicts of
interest. Thanks to the Xunta de Galicia for providing the locations of the V.
velutina nests, all anonymous people that reported nests in 2020 year and
the reviewers of the paper for the suggestions that improved the article.
References
Archer, M.E. 2006. Taxonomy, distribution and nesting biology of species of the genus
Dolichovespula (Hymenoptera, Vespidae). Entomological Science 9:
281-293. https://doi.org/10.1111/j.1479-8298.2006.00174.x
Barbet-Massin, M., Salles, J.-M., Courchamp, F. 2020. The economic cost of
control of the invasive yellow-legged Asian hornet. NeoBiota 55: 11-25. https://doi.org/10.3897/neobiota.55.38550
Beggs, J.R., Brockerhoff, E.G., Corley, J.C., Kenis, M., Masciocchi, M.,
Muller, F., Rome, Q., et al. 2011. Ecological effects and management of
invasive alien Vespidae. BioControl 56: 505-526. https://doi.org/10.1007/s10526-011-9389-z
Bessa, A.S., Carvalho, J., Gomes, A., Santarém, F. 2016. Climate and land‐use drivers of invasion: predicting the expansion of Vespa
velutina nigrithorax into the Iberian Peninsula Didham, R., Brady, S.
(eds.). Insect Conservation and Diversity 9: 27-37. https://doi.org/10.1111/icad.12140
Carvalho, J., Hipólito, D., Santarém, F., Martins, R., Gomes, A., Carmo, P.,
Rodrigues, R., et al. 2020. Patterns of Vespa velutina invasion
in Portugal using crowdsourced data. Insect
Conservation and Diversity 13: 501-507. https://doi.org/10.1111/icad.12418
Castro,
L., Pagola-Carte, S. 2010. Vespa velutina
lepeletier, 1836 (Hymenoptera: Vespidae), recolectada en la Península Ibérica. Heteropterus
Revista de Entomología 10: 193-196.
Choi, M., Kim, J., Lee, J. 2012a. Increase trend of social hymenoptera (wasps and honeybees) in urban
areas, inferred from moving‐out case by 119 rescue services in Seoul
of South Korea. Entomological Research 42: 308-319. https://doi.org/10.1111/j.1748-5967.2012.00472.x
Choi, M.B., Martin, S.J., Lee, J.W. 2012b. Distribution, spread, and
impact of the invasive hornet Vespa velutina in South Korea. Journal
of Asia-Pacific Entomology 15: 473-477. https://doi.org/10.1016/j.aspen.2011.11.004
Choi, M.B., Kim, T.G., Kwon, O. 2019. Recent Trends in Wasp Nest Removal
and Hymenoptera Stings in South Korea. Journal of Medical Entomology 56:
254-260. https://doi.org/10.1093/jme/tjy144
Diéguez-Antón, A., Escuredo, O., Seijo, M.C., Rodríguez-Flores, M.S. 2022.
Embryo, Relocation and Secondary Nests of the Invasive Species Vespa velutina
in Galicia (NW Spain). Animals 12: 2781. https://doi.org/10.3390/ani12202781
Dillane, E., Hayden, R., O’Hanlon, A., Butler, F., Harrison, S. 2022. The
first recorded occurrence of the Asian hornet (Vespa velutina) in
Ireland, genetic evidence for a continued single invasion across Europe. Journal
of Hymenoptera Research 93: 131-138. https://doi.org/10.3897/jhr.93.91209
Do, Y., Kim, J.B., Shim, J., Kim, C., Kwon, O., Choi, M.B. 2019.
Quantitative analysis of research topics and public concern on V. velutina as invasive
species in Asian and European countries. Entomological Research 49:
456-461. https://doi.org/10.1111/1748-5967.12390
Feás
Sánchez, X., Charles, R.J. 2019. Notes on the nest architecture and colony composition in winter of
the yellow-legged Asian hornet, Vespa velutina Lepeletier 1836 (Hym.:
Vespidae), in its introduced habitat in Galicia (NW Spain). Insects 10(8),
237. https://doi.org/10.3390/insects10080237
Fick, S.E.,
Hijmans, R.J. 2017. WorldClim 2: new 1km spatial
resolution climate surfaces for global land areas. International Journal of
Climatology 37 (12):4302-4315. https://doi.org/10.1002/joc.5086
Franklin, D.N., Brown, M.A., Datta, S., Cuthbertson, A.G.S., Budge, G.E.,
Keeling, M.J. 2017. Invasion dynamics of Asian hornet, Vespa velutina
(Hymenoptera: Vespidae): a case study of a commune in south-west France. Applied
Entomology and Zoology 52: 221-229. https://doi.org/10.1007/s13355-016-0470-z
Gordón-Mendoza, R., Camargo-Buitrago, I.
2015. Selección de estadísticos para la estimación de la precisión
experimental en ensayos de maíz. Agronomía Mesoamericana 26: 56-63. https://doi.org/10.15517/am.v26i1.16920
Grosso-Silva,
J.M., Maia, M. 2012. Vespa velutina
Lepeletier, 1836 (Hymenoptera, Vespidae), new species for Portugal. Arquivos
entomolóxicos 53-54.
Hartig, F. 2016. DHARMa: residual diagnostics for hierarchical
(multi-level/mixed) regression models. CRAN: Contributed Packages. https://doi.org/10.32614/CRAN.package.DHARMa.
IGE 2022. Distribución espacial de las características de la
población de Galicia por cuadrícula de 1km2. Instituto Gallego
de Estadística. Available at: https://www.ige.gal/igebdt/igeapi/csv/grid1km/2020/0/0/pob_total
[Accesed 10 May 2022].
IGN 2022a. CORINE
Land Cover 2018 (España) - CLC 2018 CC-BY 4.0. Centro Nacional de Información Geográfica (CNIG). Available
at: https://www.idee.es/csw-inspire-idee/srv/spa/catalog.search?#/metadata/spaignCLC2018
[Accesed 10 May 2022].
IGN 2022b. Modelo Digital del
Terreno de 2ª cobertura (2015-2021) con paso de malla de 200 metros (MDT200) de
España - MDT200 2015 CC-BY 4.0. Centro Nacional de
Información Geográfica (CNIG). Available at: https://centrodedescargas.cnig.es/CentroDescargas/mdt200-segunda-cobertura
[Accesed 10 May 2022].
INE 2023. Cifras oficiales de
población de los municipios españoles en aplicación de la Ley de Bases del
Régimen Local (Art. 17). Available at: https://www.ine.es/jaxiT3/Datos.htm?t=2890
[Accesed 2 January 2023].
Kennedy, P.J., Ford, S.M., Poidatz, J., Thiéry, D., Osborne, J.L. 2018.
Searching for nests of the invasive Asian hornet (Vespa velutina)
using radio-telemetry. Communications Biology 1: 88. https://doi.org/10.1038/s42003-018-0092-9
Laurino, D., Lioy, S., Carisio,
L., Manino, A., Porporato, M. 2019. Vespa velutina: An Alien Driver of Honey Bee Colony Losses. Diversity
12: 5. https://doi.org/10.3390/d12010005
Matsuura, M.,
Yamane, S. 1990. Biology of the vespine wasps.
Springer-Verlag, Berlin, Heidelberg, Germany. https://doi.org/10.1007/978-3-642-75230-8
Monceau, K., Bonnard, O., Thiéry, D. 2014. Vespa velutina: a
new invasive predator of honeybees in Europe. Journal of Pest Science
87: 1-16. https://doi.org/10.1007/s10340-013-0537-3
Monceau, K., Tourat, A., Arca, M., Bonnard, O., Arnold, G., Thiéry, D.
2017. Daily and Seasonal Extranidal Behaviour Variations in the Invasive
Yellow-Legged Hornet, Vespa velutina Lepeletier (Hymenoptera:
Vespidae). Journal of Insect Behavior 30: 220-230. https://doi.org/10.1007/s10905-017-9607-3
Pazos, T., Álvarez-Figueiró, P., Cortés-Vázquez, J.A., Jácome, M.A.,
Servia, M.J. 2022. Of Fears and Budgets: Strategies of Control in Vespa velutina
Invasion and Lessons for Best Management Practices. Environmental Management
70: 605-617. https://doi.org/10.1007/s00267-022-01690-z
Perrard, A., Haxaire, J., Rortais, A., Villemant, C. 2009. Observations on
the colony activity of the Asian hornet Vespa velutina Lepeletier 1836
(Hymenoptera: Vespidae: Vespinae) in France. Annales de la Société
entomologique de France (N.S.) 45: 119-127. https://doi.org/10.1080/00379271.2009.10697595
Poidatz, J., Monceau, K., Bonnard, O., Thiéry, D. 2018. Activity rhythm and action range of workers of the invasive hornet
predator of honeybees Vespa velutina, measured by radio frequency
identification tags. Ecology and Evolution 8: 7588-7598. https://doi.org/10.1002/ece3.4182
Porporato, M., Manino, A., Laurino, D., Demichelis, S. 2014. Vespa velutina
Lepeletier (Hymenoptera Vespidae): a first assessment two years after its
arrival in Italy. Redia 97: 189-194.
R Core Team 2022. R: A language and environment for statistical computing. R
Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Robinet, C., Suppo, C., Darrouzet, E. 2017. Rapid spread of the invasive
yellow‐legged hornet in F
rance: the role of human‐mediated dispersal and the effects of control measures Diekötter, T.
(ed.). Journal of Applied Ecology 54: 205-215. https://doi.org/10.1111/1365-2664.12724
Rodríguez-Flores, M.S., Seijo-Rodríguez, A., Escuredo, O., Seijo-Coello, M.D.C.
2019. Spreading of Vespa velutina in northwestern Spain:
influence of elevation and meteorological factors and effect of bait trapping
on target and non-target living organisms. Journal of Pest Science 92:
557-565. https://doi.org/10.1007/s10340-018-1042-5
Rojas-Nossa, S.V., Novoa, N., Serrano, A., Calviño-Cancela, M. 2018. Performance of baited traps used as control tools for the invasive
hornet Vespa velutina and their impact on non-target insects. Apidologie
49: 872-885. https://doi.org/10.1007/s13592-018-0612-0
Rojas-Nossa, S.V., Gil, N.,
Mato, S., Garrido, J. 2021. Vespa velutina: traits and impacts of a successful invasive alien
species. Ecosistemas 30: 1-10.
https://doi.org/10.7818/ECOS.2159
Rojas-Nossa, S.V., Álvarez, P.,
Garrido, J., Calviño-Cancela, M. 2022. Method for Nest
Detection of the Yellow-Legged Hornet in High Density Areas. Frontiers in
Insect Science 2: 851010. https://doi.org/10.3389/finsc.2022.851010
Rome, Q., Muller, F.J., Touret‐Alby, A., Darrouzet, E.,
Perrard, A., Villemant, C. 2015. Caste differentiation and seasonal changes in Vespa
velutina (Hym.: Vespidae) colonies in its introduced range. Journal of
Applied Entomology 139: 771-782. https://doi.org/10.1111/jen.12210
Rome, Q., Perrard, A., Muller, F., Fontaine, C., Quilès, A., Zuccon, D.,
Villemant, C. 2021. Not just honeybees: predatory habits of Vespa velutina
(Hymenoptera: Vespidae) in France. Annales de la Société
entomologique de France (N.S.) 57: 1-11. https://doi.org/10.1080/00379271.2020.1867005
Venables,
W.N., Ripley, B.D. 2002. Random
and Mixed Effects. In Modern Applied Statistics with S, Statistics and
Computing, pp. 271-300. Springer New York, NY, USA. https://doi.org/10.1007/978-0-387-21706-2_10
Villemant, C., Barbet-Massin, M., Perrard, A., Muller, F., Gargominy, O.,
Jiguet, F., Rome, Q. 2011a. Predicting the invasion risk by the alien
bee-hawking Yellow-legged hornet Vespa velutina nigrithorax
across Europe and other continents with niche models. Biological
Conservation 144: 2142-2150. https://doi.org/10.1016/j.biocon.2011.04.009
Villemant, C., Muller, F., Haubois, S., Perrard, A., Darrouzet, E., Rome, Q.
2011b. Bilan des travaux (MNHN et IRBI) sur l’invasion en France de Vespa
velutina, le frelon asiatique prédateur d’abeilles. Proceedings
of the Journée Scientifique Apicole–11 February 3-12.
Williams, P. 1988. Social wasps (Hym., Vespidae) from the Kashmir Himalaya. Entomologist's
Monthly Magazine, 124(1488–1491):149–152.
Wood, S.N. 2017. Generalized Additive Models: An Introduction with R
(2nd edition). Chapman and Hall/CRC, New York.
Xunta de
Galicia 2020a. RESOLUCIÓN de 16 de enero
de 2020, de la Secretaría General Técnica de la Consellería de Presidencia,
Administraciones Públicas y Justicia, por la que se da publicidad al encargo a
la Empresa Pública de Servicios Agrarios Gallegos, S.A. (Seaga) para la tramitación
y ejecución de los trabajos de retirada y destrucción de nidos incluidos en el
Programa gallego de vigilancia y control frente a la avispa asiática (avispa
velutina). Avaliable at: https://www.xunta.gal/dog/Publicados/2020/20200117/AnuncioG0244-160120-0003_es.html
[Accesed 5 May 2023].
Xunta de Galicia 2020b. RESOLUCIÓN
de 16 de septiembre de 2020 por la que se da publicidad de las personas
beneficiarias de las ayudas concedidas al amparo de la Orden de 20 de mayo de
2020 por la que se establece el régimen de ayudas a la apicultura y se convocan
para el año 2020. Avaliable at: https://www.xunta.gal/dog/Publicados/2020/20200925/AnuncioG0426-170920-0002_es.html
[Accesed 5 May 2023].
Zeileis, A., Kleiber, C., Jackman, S. 2008. Regression Models for Count
Data. R. Journal of Statistical Software 27: 1-25. https://doi.org/10.18637/jss.v027.i08
Appendix / Anexo
Table A1. Exponentiated Coefficients from the
Zero-Inflated Model.
Tabla A1. Coeficientes exponenciales del modelo
cero inflado.
|
Variable
|
Count Component
|
Zero-Inflation Component
|
|
Intercept
|
23.117
|
16.458
|
|
Population
(Nº people)
|
10.001
|
0.9459
|
|
Altitude
(m)
|
0.9969
|
10.020
|
|
Agricultural land cover (Cell %)
|
28.402
|
0.5157
|
|
Urban land cover (Cell %)
|
97.332
|
0.2835
|