Introducción
Servicios
ecosistémicos suministrados por la avifauna
El contexto actual de
cambio global y crisis climática está provocado, entre otros factores, por una
deficiente integración de las actividades humanas en los ecosistemas (Liu et al. 2007). La
producción agraria y forestal intensiva se soporta mediante técnicas de
fertilización y control de plagas poco sostenibles desde el punto de vista de
la biodiversidad y la gestión de los recursos naturales (Bommarco et al. 2013). Una
manera de avanzar hacia sistemas agroforestales más sostenibles es incorporar
los servicios que proporciona la biodiversidad en los sistemas de producción (Landis 2017). Por ejemplo, los métodos de
control biológico de plagas basados en depredadores naturales, servicio de
regulación según la clasificación del Millennium Ecosystem Assessment (2005), permiten reducir la aplicación de plaguicidas,
la contaminación que producen en alimentos, suelos, sistemas acuáticos y
cadenas tróficas, y reducir el impacto de los plaguicidas sobre la salud humana
y la biodiversidad (Monteagudo et al. 2023).
Otra
manera de avanzar hacia sistemas agroforestales más sostenibles es reconducir la política forestal de repoblaciones masivas, a veces poco
exitosas, hacia políticas de facilitación o aceleración de la regeneración
natural forestal incorporando los servicios de dispersión de semillas que
proporciona la fauna.
Por ejemplo, si conocemos
los dispersores de bellotas y su comportamiento dispersivo en distintos
contextos ambientales, podremos facilitar
estos procesos y amplificarlos en el paisaje,
generando un incremento de la diversidad biológica y un importante ahorro
económico (Hougner et al. 2006). Este servicio suele enclavarse dentro de los servicios
de soporte (o de apoyo), según la clasificación del Millennium Ecosystem Assessment (2005), con implicaciones en servicios de
aprovisionamiento y culturales. Desafortunadamente, en muchos casos
desconocemos la identidad de las especies más importantes desde un punto de
vista funcional (aquellas con capacidad para proporcionar más servicios ecosistémicos)
y las características de sus funciones en los ecosistemas. Esto limita nuestra capacidad para incorporar la
biodiversidad en los sistemas de gestión y desarrollar sistemas agroforestales
más sostenibles, es decir, con más biodiversidad y más resilientes a las
perturbaciones.
La avifauna es un grupo
de vertebrados singular. Su capacidad de vuelo les permite sortear barreras
físicas mejor que otros vertebrados y desplazarse entre teselas que están
alejadas a escala del paisaje, una escala espacial relevante para organizar
sistemas agrarios más sostenibles (Boesing et al. 2017). La
avifauna es un componente esencial de las “high mobile link species”,
con capacidad para realizar funciones variadas y conectar funcionalmente las
teselas de los ecosistemas (Lundberg y Moberg 2003). Aunque se han descrito servicios ecosistémicos de soporte,
aprovisionamiento, regulación y culturales proporcionados por la avifauna (Whelan et al. 2008), queda
mucho trabajo por hacer para conseguir una cuantificación y caracterización
adecuadas de los mismos, que permita incorporar la avifauna a los sistemas de
gestión y producción para hacerlos más sostenibles (Rebollo et al. 2019; García et al. 2021, 2024;
González-Varo
et al. 2023).
En este artículo sintetizamos e integramos los trabajos
realizados por el equipo de investigación combinando el fototrampeo con
técnicas de seguimiento de la actividad de la avifauna. Por un lado, combinamos
el fototrampeo y el seguimiento con radiotransmisores (emisores de
radiofrecuencia) para analizar el papel de los córvidos en la dispersión de
semillas grandes (principalmente bellotas) que dependen de la sinzoocoria para
su dispersión. La sinzoocoria es un mutualismo en el que los animales
transportan voluntariamente las bellotas y las almacenan para su posterior
consumo (García et al. 2010; Farwig y Berens 2012).
Algunas especies de córvidos, como el arrendajo euroasiático (Garrulus
glandarius) y la urraca común (Pica pica), intervienen decisivamente
en los procesos de regeneración natural de los bosques de Quercus (Pesendorfer et al.
2016; Gómez et al. 2019), un género
de plantas leñosas con elevada importancia ecológica, económica y social en
áreas extensas de Iberia y del hemisferio norte (Gil-Pelegrín et al. 2017). En Iberia hay
una elevada variedad de especies de córvidos y desconocemos el papel de las
diferentes especies en la dispersión de quercíneas u otras plantas en distintos
contextos ambientales. También desconocemos detalles fundamentales de su
actividad dispersiva como la distancia de dispersión, las características de
los lugares de almacenamiento o los procesos de recuperación-consumo de
bellotas en los almacenes.
Por otro lado,
combinamos técnicas de fototrampeo y de seguimiento con emisores GPS (satélite)
para estudiar el papel de las aves rapaces en la regulación de plagas
agrícolas. Tomamos como modelo el abejero europeo (Pernis apivorus, de
aquí en adelante, abejero) y la avispa asiática (Vespa velutina nigrithorax,
de aquí en adelante, velutina), una avispa exótica invasora que genera
importantes problemas económicos, ecológicos y sanitarios a nivel europeo (Rojas-Nossa y Calviño-Cancela 2020). La
velutina se introdujo accidentalmente en Francia desde China en 2004,
expandiéndose rápidamente por el continente europeo (Arca et al. 2015; Budge et al. 2017). En
2014, la Unión Europea la incluyó en la lista de especies exóticas invasoras
que causan mayores daños (UE1143/2014). Pocos estudios han
analizado la capacidad de las aves rapaces para controlar plagas agrícolas y
formar parte de programas de lucha integral contra las plagas (Integrated Pest
Management, IPM) (Monteagudo et al. 2023). En
2018 iniciamos una línea de investigación sobre el abejero europeo, una de las
rapaces diurnas europeas más desconocidas (Hagemeijer y Blair 1997). Es
una rapaz migratoria que se reproduce en Europa y Asia occidental e inverna en
África subsahariana (Gamauf y Haring 2004). Su
estancia en Europa es corta, principalmente de mayo a agosto, ambos inclusive.
Durante el periodo reproductor se comporta como un depredador especialista que
consume principalmente véspidos eusociales (Cramp y Simmons 1980) y
podría incluir la velutina en su dieta. Ataca los avisperos y lleva al nido
fragmentos de panales cargados con larvas y pupas con las que alimenta a las
crías.
Objetivo
Objetivo general
El objetivo general de este trabajo es mostrar el uso de técnicas
de fototrampeo combinadas con técnicas de seguimiento con emisores para
estudiar dos servicios ecosistémicos de la avifauna: la dispersión de semillas
grandes y la regulación de plagas. Es un trabajo de revisión donde se
sintetizan e integran trabajos realizados por el grupo de investigación, aunque
también se presentan resultados inéditos sobre la interferencia del fototrampeo
en el comportamiento de las especies diana y sobre los fallos técnicos
detectados en nuestros estudios.
Objetivos específicos
Mostrar la eficacia del fototrampeo
y las técnicas de seguimiento con emisores para identificar la avifauna que
lleva a cabo servicios ecosistémicos de control de plagas y dispersión de
semillas, y caracterizar el servicio. Este objetivo incluye identificar las
especies de aves implicadas y el tipo de individuos (sexo, edad) que
proporcionan el servicio. Además, cuantificar el servicio y caracterizar su
escala espacial y temporal mediante el seguimiento con emisores de aves y
semillas. Este análisis contribuye a resolver aspectos relacionados con la
identificación precisa de los proveedores de servicios ecosistémicos y la
comprensión de la dinámica temporal y espacial de estos servicios.
Analizar la interferencia del fototrampeo en el
comportamiento de la avifauna diana. Se examinan los cambios en el
comportamiento de las aves en presencia de las cámaras de fototrampeo y se
determinan los tiempos de habituación a las cámaras. Esto aporta conocimiento
clave sobre las limitaciones metodológicas del fototrampeo que pueden influir
en la interpretación de los resultados obtenidos.
Analizar los problemas técnicos de las cámaras de
fototrampeo. Se tipifican y cuantifican los principales fallos técnicos de los
equipos, discutiendo las posibles soluciones. Esta evaluación contribuye a
mejorar la fiabilidad de las técnicas utilizadas y optimizar su aplicación en
futuros estudios.
Con toda esta información, realizamos una síntesis final
sobre la contribución del fototrampeo al estudio de los servicios ecosistémicos
suministrados por la avifauna y las sinergias entre el fototrampeo y las
tecnologías de seguimiento con emisores. Evaluamos no
solo la eficacia del fototrampeo para identificar especies proveedoras de
servicios ecosistémicos, sino también las interferencias del fototrampeo en el comportamiento
de las especies diana y los problemas técnicos asociados al fototrampeo.
Además, caracterizamos los servicios ecosistémicos combinando el fototrampeo
con técnicas de seguimiento por radiofrecuencia o GPS. Esta combinación de
técnicas representa un avance metodológico significativo en el estudio de las funciones
de las aves en los ecosistemas y proporciona una base sólida para futuras
investigaciones sobre sus servicios ecosistémicos.
Líneas de investigación desarrolladas por el equipo y técnicas de fototrampeo y de seguimiento con
emisores utilizadas
Interacción
córvido-bellota
En 2009 iniciamos una línea de investigación sobre
dispersión de bellotas y otras semillas grandes por córvidos y, en menor medida, ratones, que continúa en la actualidad (Pérez-Camacho et al. 2023). Las áreas de estudio estaban ubicadas en Uceda
(Guadalajara) y Alcalá de Henares (Madrid), en un ecosistema agroforestal
boscoso y otro adehesado, respectivamente, ambos dominados por encina y con
clima mediterráneo continental (Martínez-Baroja et al. 2019, 2021; Pérez-Camacho
et al. 2023). El fototrampeo fue una herramienta esencial y los
modelos de cámaras evolucionaron a lo largo del tiempo: Bushnell Trailscout 2.1
MP, Moultrie MCG-12634, Browning Dark Ops HD Elite y, finalmente, Browning Spec
Ops (Tabla 1). Las cámaras se instalaron en las
proximidades de comederos, donde depositábamos las bellotas, para grabar en
vídeo su remoción por dispersores y consumidores (Fig. 1). Se configuraron para grabar
un vídeo de la calidad más alta posible (de 240p a 1080p según modelo) de un
minuto de duración cada vez que detectaban movimiento, y el menor tiempo
posible de pausa entre vídeos. Estaban instaladas entre 1.2 y 2 m de distancia
al comedero, en el suelo o en árboles a poca altura (máx. 3 m), lo que
permitió comprobar fácil y regularmente el funcionamiento de las cámaras y los
accesorios, por ejemplo, las baterías (Fig. 2).
Para el seguimiento posterior de las bellotas removidas por
los córvidos, se insertó dentro de la bellota un emisor de radiofrecuencia
(PIP5 de Biotrack) con la antena enrollada sobre sí mismo siguiendo la
metodología de Pons
y Pausas (2007) (Fig. 3). Las bellotas con
emisor tenían un aspecto similar a las bellotas normales y fueron removidas de
los comederos por los córvidos de manera similar. Las bellotas dispersadas por
los córvidos se localizaron posteriormente mediante un receptor de radiofrecuencia
con una antena unidireccional Yagi (Biotrack). Para la localización exacta se
usó un detector de metales de mano (White’s Auto-Scan Personal Search Detector)
(Castro et al. 2017; Martínez-Baroja
et al. 2019, 2021; Pérez-Camacho et al. 2023). Para
identificar en los vídeos a los individuos que utilizaban los comederos,
algunos ejemplares fueron capturados (33 adultos y
136 pollos), marcados con anillas de colores,
y los adultos fueron sexados mediante análisis genéticos (19 hembras y 14 machos).
Esto permitió asignar a individuos concretos (según sexo, estado
reproductor, etc.) los comportamientos grabados, así como cuantificar las
bellotas dispersadas y conocer su patrón espacial de dispersión (Martínez-Baroja et al. 2019, 2021).
Tabla 1.
Características de las cámaras de fototrampeo utilizadas en los estudios
analizados en este artículo.
Table 1.
Characteristics of the camera traps used in the studies analysed in this
article.
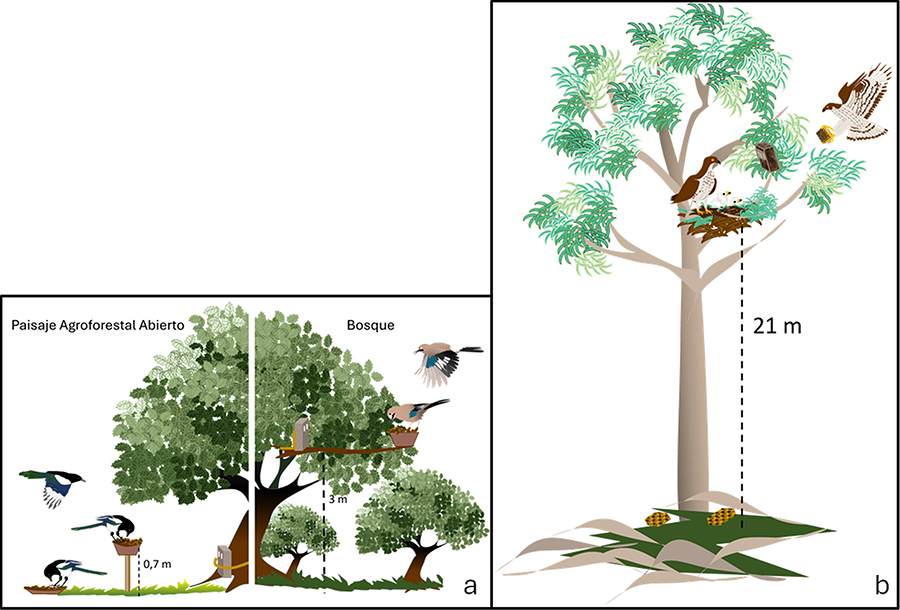
Figura 1. Este estudio evalúa sistemas de fototrampeo
instalados en circunstancias ambientales y de trabajo contrastadas. En el caso
de la interacción córvido-bellota (a), las cámaras fueron instaladas en lugares
fácilmente accesibles con el objetivo de registrar el comportamiento dispersivo
de bellotas por los córvidos en la copa de encinas, en comederos situados sobre
el suelo, en comederos situados al final de barras verticales a 70 cm sobre el
suelo, o en comederos situados en ramas de árboles situadas a 3 m de altura. En
el caso de la interacción abejero-velutina (b), las cámaras fueron instaladas
en las proximidades de los nidos de abejero, en lugares muy poco accesibles
situados normalmente en eucaliptos a una altura media de 21 m sobre el suelo.
Figure 1. This
study assesses camera-trapping systems deployed under diverse environmental
conditions and operational settings. For the corvid-acorn interaction (a),
cameras were installed in easily accessible locations to capture the acorn
dispersal behavior of corvids, including behaviours in the crowns of oak trees,
feeders placed on the ground, feeders mounted on vertical poles 70 cm above the
ground, and feeders positioned on tree branches at a height of 3 meters. For
the honey buzzard-velutina interaction (b), cameras were installed near honey
buzzard nests, in highly inaccessible locations, typically within eucalyptus
trees at an average height of 21 meters above the ground.

Figura 2. Montajes de las cámaras de
fototrampeo en los distintos experimentos de la interacción córvido-bellota: a)
comedero para arrendajo con cámara de fototrampeo situado en la copa de una
encina a unos 3 m de altura sobre el suelo; b) cámara de fototrampeo enfocando
hacia una encina con bellotas (sin comedero); c) comedero para urraca con
cámara de fototrampeo situado sobre el suelo; d) comedero para urraca con
cámara de fototrampeo situado al final de una barra vertical de unos 70 cm
sobre el suelo. Las cámaras más modernas en b, c y d tienen conectadas baterías
externas.
Figure 2.
Camera trap setups used in various experiments investigating the corvid-acorn
interaction: a) a jay feeder equipped with a camera trap mounted in the crown
of an oak tree, approximately 3 meters above the ground; b) a camera trap aimed
at an oak tree containing acorns (without a feeder); c) a magpie feeder with a
camera trap placed on the ground; d) a magpie feeder with a camera trap
installed at the end of a vertical pole, approximately 70 cm above the ground.
The cameras used in setups b), c), and d) are connected to external batteries.

Figura 3. Inserción del radiotransmisor y su
antena en el interior de la bellota en los estudios de dispersión por córvidos.
El aspecto externo, la consistencia y el peso de la bellota con radiotransmisor
son similares al de una bellota normal.
Figure 3.
Insertion of a radio transmitter and its antenna into an acorn for studies on
dispersal by corvids. The external appearance, consistency, and weight of the
acorn containing the radio transmitter remain similar to those of a typical
acorn.
Interacción abejero-velutina
Para conocer si el abejero depredaba sobre la velutina,
localizar y conocer el tipo de avisperos atacados, y cuantificar el grado de
destrucción provocado en los mismos, en el periodo 2018-2022 instalamos cámaras
de fototrampeo modelo Browning Spec Ops en 33 nidos de abejero estudiando su
dieta a través del registro fotográfico (Fig. 1).
El área de estudio estaba ubicada en la península del
Morrazo (Pontevedra), en un ecosistema agroforestal boscoso con eucaliptos de
repoblación y manchas de bosque autóctono en clima atlántico (Martín‐Ávila
et al. 2024). El abejero construyó
sus nidos sobre grandes árboles a una altura media de 21 m (rango 7.4 – 34.8
m). En los tres primeros años instalamos cámaras modelo Browning Spec Ops en
los nidos a principios de julio, una vez que los pollos eran capaces de
termorregular y podían mantener su temperatura corporal en ausencia de los
adultos, a partir de 14 días de edad (Tabla 1).
Los dos últimos años instalamos cámaras modelo Browning Spec Ops Edge en abril,
antes de que los abejeros adultos llegaran a Europa, en los nidos ocupados la
temporada de reproducción anterior. Esto permitió obtener el registro
fotográfico de todas las etapas de la reproducción (reparación del nido,
incubación, crianza de los pollos). En aquellos casos en los que los abejeros
ocuparon otros nidos, instalábamos las cámaras a principios de julio, tal y
como lo habíamos realizado en años anteriores. Las cámaras estaban instaladas
en las proximidades del nido, a una altura elevada y a una distancia de 1 a 2
metros del nido (Figs. 1 y 4). Son lugares poco accesibles excepto con
sistemas de escalada especializados. Debido a la poca accesibilidad y a las
molestias que se ocasionarían a los individuos reproductores, la comprobación
de las cámaras y accesorios (p.e. baterías) sólo podía realizarse en contadas
ocasiones, por lo que fueron necesarios equipos de fototrampeo con elevada
autonomía energética y fiabilidad.
Para localizar los avisperos atacados por los abejeros,
durante dos años capturamos 11 abejeros adultos y les instalamos emisores GPS
con señal registrada por satélite (modelo Ornitrac-20 de Ornitela®) que
permitían el registro digital de la posición de los individuos y, por tanto, el
análisis geoespacial de sus movimientos mediante softwares especializados (SIG)
(Fig. 5). En los 17 nidos donde teníamos adultos
de abejero con emisores GPS, además de la cámara Browning Spec Ops, instalamos
una segunda cámara Spromise S378 4G que enviaba fotos al ordenador en tiempo
real cada vez que detectaba la llegada de adultos al nido con presas (Fig. 4; Tabla 1).
Cuando detectábamos que el abejero había llevado un panal de velutina al nido,
podíamos rastrear las posiciones del adulto y localizar y monitorear los
avisperos atacados en el campo.
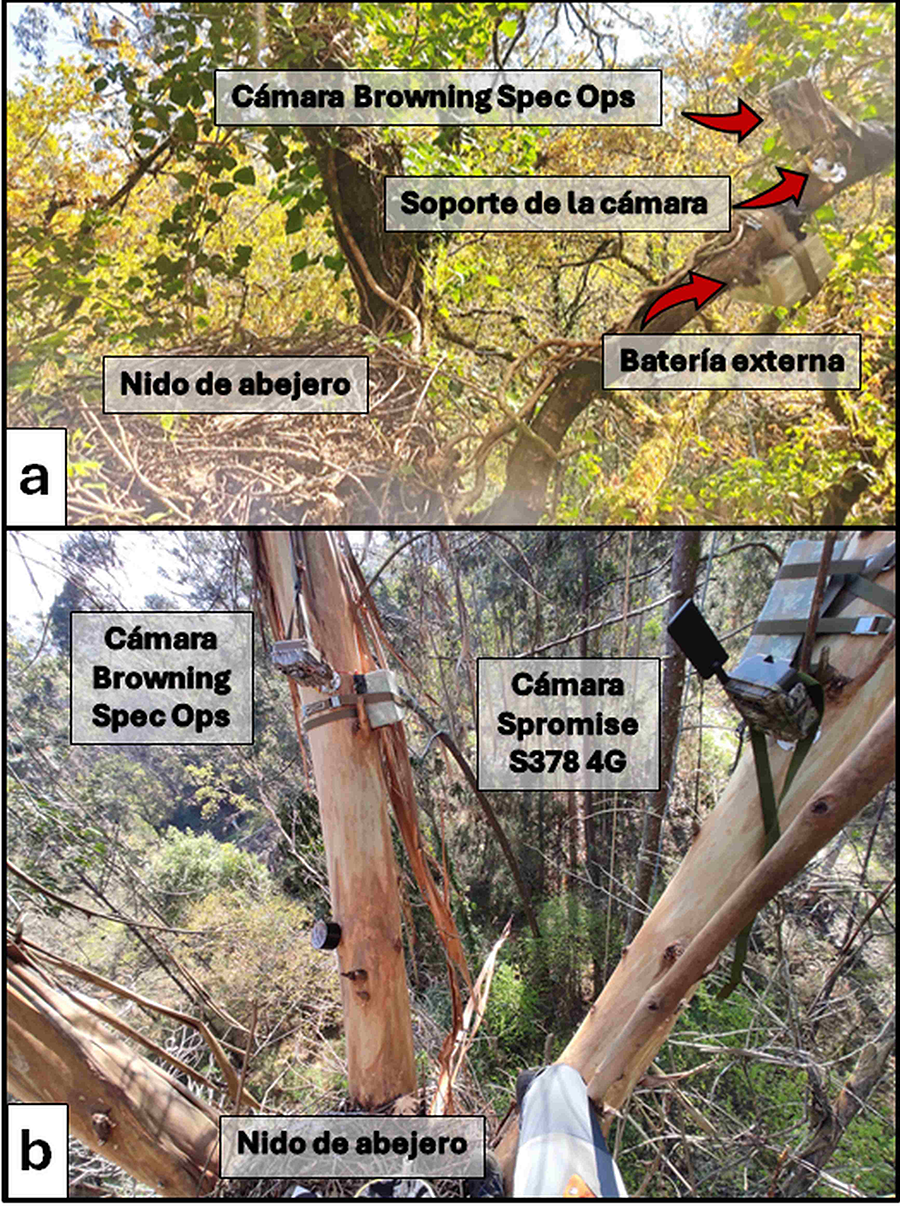
Figura 4. Montaje de
las cámaras de fototrampeo y baterías externas en las proximidades de los nidos
de abejero europeo: a) montaje con una sola cámara Browning Spec Ops; y b)
montaje con dos cámaras, una Browning Spec Ops y una Spromise S378 4G (en esta
última puede apreciarse la antena que permite enviar las fotos directamente al
ordenador).
Figure 4.
Installation of camera traps and external batteries near European honey-buzzard
nests: a) an installation with a single Browning Spec Ops camera; and b) an
installation with two cameras, including a Browning Spec Ops and a Spromise
S378 4G (which features an antenna allowing direct photo transmission to a
computer).

Figura 5. Hembra de
abejero fotografiada en el nido alimentando a su pollo con larvas de Vespa
velutina, en la que se aprecia el emisor GPS (satélite) sujeto a su espalda
mediante un arnés adaptado a su tamaño. Este emisor tiene un pequeño panel
solar que permite la recarga de la energía que necesita. La foto está tomada
con la cámara Browning Spec Ops. En la parte superior derecha se muestra una
imagen del emisor GPS completo.
Figure 5. A female honey-buzzard photographed at the nest as she feeds her
chick with larvae of Vespa velutina. The satellite GPS transmitter is
visible on her back, secured with a harness tailored to her size. This
transmitter includes a small solar panel that recharges it using solar energy.
The photograph was taken using a Browning Spec Ops camera. A complete image of
the GPS transmitter appears in the upper right corner.
Eficacia del fototrampeo para
identificar la avifauna que realiza los servicios ecosistémicos y caracterizar
los servicios
Interacción córvido-bellota
En un estudio llevado a cabo en un
bosque mixto de encina y quejigo, se instalaron tres cámaras Bushnell Trail
Scout 2.1 MP en las copas de las encinas, registrando más de 600 vídeos que
documentaron la visita de aves a los comederos (Pérez-Camacho et al. 2023). Se identificaron seis especies de aves en total, aunque
el 99% de los registros correspondieron a arrendajos, que fueron la única
especie observada manipulando, consumiendo y retirando bellotas (Vídeos S1 y S2). Se identificaron
cuatro comportamientos de los arrendajos. 1) Selección de bellotas: En el 25%
de los vídeos, los arrendajos seleccionaron bellotas tras manipularlas
repetidamente con el pico, dejándolas caer y eligiendo finalmente una. 2) Consumo
en el sitio: En el 4% de los casos, las bellotas seleccionadas fueron
consumidas en el lugar. 3) Transporte de bellotas: En el 35% de los vídeos se
observó a los arrendajos volando con bellotas intactas alejándose del comedero.
4) Otros comportamientos (34% restante) (Tabla 2). De los vuelos, el 66% implicó una sola bellota en el
pico, el 3% una bellota en la bolsa gular, y el 31% el transporte simultáneo de
dos bellotas (una en la bolsa gular y otra en el pico).
Tabla 2.
Comportamiento de los arrendajos (Garrulus glandarius) en los comederos.
Table 2. Eurasian jay (Garrulus glandarius) behavior at feeders.
La combinación de fototrampeo y el
uso de radiotransmisores insertados en las bellotas permitió determinar que los
arrendajos almacenaron el 64.2% de las bellotas (192 de un total de 299
bellotas radiomarcadas), mientras que el 35.8% restante (107 bellotas) fueron
consumidas en el comedero o en sus inmediaciones. Este uso complementario de
cámaras y radiotransmisores confirmó el comportamiento de almacenamiento
disperso de los arrendajos, caracterizado por depositar una bellota en cada
sitio de almacenamiento y exhibir un comportamiento compulsivo de acopio (Pérez-Camacho et al. 2023). Además, se identificaron variaciones en el proceso de
almacenamiento dependiendo de la posición de los árboles madre y la estructura
del hábitat, sugiriendo ligeras diferencias en el comportamiento individual de
los arrendajos.
En formaciones forestales abiertas
con encinas y quejigos dispersos, se instalaron 14 cámaras Browning Dark Ops HD
Elite que proporcionaron más de 400 vídeos que mostraban aves cogiendo bellotas
de la copa de las encinas, el 65% de las aves fueron urracas. También seis
cámaras del mismo tipo proporcionaron más de 200 vídeos que mostraban aves y
mamíferos cogiendo bellotas del suelo bajo las encinas, el 48% de los cuales
fueron de urraca (Martínez-Baroja
et al. 2019).
Posteriormente, anillando urracas, grabando en vídeo con 14 cámaras más de 7500
eventos de remoción de bellotas en comederos y colocando radiotransmisores a
337 bellotas, determinamos que (1) la urraca era la única especie de córvido
que almacenó bellotas, almacenando el 45% de la producción anual en escondites
individuales en el suelo; (2) las parejas reproductoras de urraca, y
especialmente los machos, fueron los principales dispersores de bellotas; y (3)
cada urraca reproductora almacenó entre 169 y 1372 bellotas durante el periodo
de dispersión (6 semanas). El uso combinado de estas dos técnicas permitió
demostrar que la urraca es una dispersora masiva y efectiva de bellotas en
sistemas forestales abiertos (Martínez-Baroja
et al. 2019).
Además, la obtención de
información espacial adicional, como la posición de los nidos de urraca y las
características espaciales de sus hábitats, demostró que el almacenamiento de
bellotas es consecuencia de una toma de decisiones jerárquica y estructurada.
Los individuos eligen los sitios de almacenamiento guiados primero por la
territorialidad y luego por sus preferencias de ubicación a diferentes escalas
espaciales (Martínez‐Baroja et al. 2021). Este comportamiento
influye en la estructura espacial de la población de plantas de quercíneas
dispersadas. El fototrampeo permitió observar el proceso de almacenamiento de
algunas bellotas por las urracas en distintos sustratos tales como tierra removida,
hojarasca, suelo pedregoso, etc. Las urracas seleccionan el sitio de
almacenamiento, generalmente tras varios intentos, introducen la bellota en el
suelo por su parte afilada y martillean con el pico en su parte más ancha.
Posteriormente ocultan la bellota con el material circundante: tierra, hojas
secas, hojas de herbáceas arrancadas por ellas, o piedras, en ocasiones muy
grandes (hasta 696 g) para el peso de las urracas (184 g hembras y 201 g machos).
Esporádicamente, también grabamos la recuperación de bellotas almacenadas por
la urraca (Vídeos S3 a S14).
Miembros de nuestro equipo de investigación de la Universidad de Granada han
estudiado la dispersión de nueces por urraca también con estas técnicas (Castro et al. 2017; Molina-Morales
et al. 2019, 2020, 2023).
Interacción abejero-velutina
Las 33 cámaras Browning Spec Ops
instaladas en los nidos de abejero realizaron un total de 996 476 fotos (rango
2089 – 99 107 por cámara). El análisis del registro fotográfico permitió
identificar un total de 5893 presas llevadas al nido por los adultos (rango 0 –
419 presas por cámara), siendo el mayor número de presas del abejero
registradas hasta la fecha (Rebollo et al. 2023; Martín‐Ávila
et al. 2024). El 82% de las presas fueron trozos de panales de
véspidos y, en menor medida, de otros himenópteros. La velutina fue consumida
por todas las parejas de abejero estudiadas, siendo la segunda presa más
importante en el área de estudio (el 24.2% de las presas llevadas al nido
fueron panales de velutina, Martín‐Ávila et al. 2024) (Vídeos S15 y S16). No detectamos ningún caso de depredación
sobre la abeja doméstica (Apis mellifera). Las fotos permitieron
diferenciar panales con celdillas grandes y panales con celdillas pequeñas,
pero fue necesario recoger muestras de ambos tipos de panales en el nido del
abejero y sus inmediaciones para identificar los panales hasta nivel específico
en el laboratorio. El 99% de los panales con celdillas grandes pertenecían a
velutina y el 99.2% de los panales de celdillas pequeñas a avispas comunes (Vespula
vulgaris y V. germánica). El 16.7% de las presas fueron pequeños
vertebrados (reptiles, aves, anfibios y mamíferos), siendo
su presencia infravalorada en los estudios anteriores
basados en el análisis de restos de presas (Martín‐Ávila
et al. 2024). El registro fotográfico permitió estimar el número y
tamaño de los panales transportados al nido y estimar
aproximadamente el número de avisperos de
velutina destruidos por el abejero (Rebollo et al. 2023).
Para ello se tuvo en cuenta el número y el tamaño de los panales en los
avisperos de velutina durante la estancia del abejero en Europa (Rome et al. 2015). Por tanto, el fototrampeo fue
un procedimiento eficaz y poco sesgado de estudio de la dieta del abejero, como
se demostró también en estudios de la dieta de otras rapaces (García-Salgado et al. 2015). Debido al
notable dimorfismo sexual en el plumaje de los abejeros, el registro
fotográfico está permitiendo analizar por separado las presas aportadas por el
macho y la hembra, y conocer el papel de ambos sexos durante el periodo
reproductor.
La utilización simultánea
de cámaras Spromise S378 4G instaladas en 17 nidos de abejero y de emisores GPS
(satélite) instalados en los adultos de abejero nos permitió conocer la
localización muy aproximada de 572 avisperos atacados (206 de velutina y 366 de
otros véspidos). De ellos pudimos localizar físicamente en el campo un total de
307 avisperos atacados. El 86% de los avisperos de velutina fueron avisperos
primarios (avisperos anteriores al avispero secundario o definitivo) situados
en el suelo y mayoritariamente dentro de manchas forestales (Vídeo S17).
El grado de destrucción inferido fue completo en el 89.25% de los casos. Por
tanto, el fototrampeo junto con tecnologías de seguimiento GPS de los abejeros
adultos permitió abordar los tres objetivos propuestos en nuestra
investigación. Otras investigaciones complementarias, no basadas en técnicas de
fototrampeo, permitieron contestar a otra pregunta clave: si esta presión de
depredación tiene un efecto demográfico sobre la velutina y provoca una
reducción en el número de obreras y avisperos alrededor de los nidos de abejero
(Martín-Ávila et al. 2025).
Cambios en el comportamiento de la avifauna en presencia
de las cámaras de fototrampeo
Interacción córvido-bellota
Los cambios en el comportamiento
de los córvidos en presencia del equipo cámara-comedero fueron dependientes de
la especie de córvido. En los vídeos se aprecia que los arrendajos modificaron
poco su comportamiento. En algún caso se observó que miraban detenidamente la
cámara, a veces desde muy cerca, y erizaban las plumas del píleo, como señal de
nerviosismo, pero su comportamiento en los comederos fue tranquilo en la
mayoría de las grabaciones. Utilizamos distintos tipos de comedero: metálicos,
de mimbre, etc. y con distintas formas y colores, con pintura de camuflaje y
sin ella, sin observar diferencias apreciables de comportamiento.
Sin embargo, la urraca es una
especie neófoba que se mostró mucho más recelosa con el equipo cámara-comedero.
El comedero fue el elemento que produjo más perturbaciones en el comportamiento
de las urracas. Así, el tipo de comedero fue clave para nuestro estudio.
Ciertos tipos de comederos no fueron visitados ninguna vez por las urracas,
otros solo ocasionalmente y otros fueron usados con diferencias de
comportamiento entre individuos (Figura S3 en Martínez-Baroja et al. 2019). Los comederos que mejor
funcionaron con las urracas fueron los situados sobre el suelo, que consistieron
en un plato circular de plástico de 20 cm de diámetro semienterrado con un
agujero en el centro y fijado al suelo con un clavo, y que los utilizamos sólo
en experimentos concretos porque también son accesibles a ratones de campo y
conejos. Tras ellos, los comederos situados a 70 cm del suelo (en el extremo de
una barra vertical que nos permitía excluir a ambos mamíferos; Figs. 1 y 2). Estos comederos fueron fabricados con un colador
doméstico semiesférico de 17 cm de diámetro con malla metálica fina para que
drenase el agua de lluvia y fueron pintados con colores del medio para
camuflarlos. Aunque una buena parte de las
urracas se acostumbró a estos comederos tras un tiempo, otra fracción de la
población presentó un comportamiento nervioso al retirar las bellotas,
posándose en el comedero tras varios intentos de subir a él desde el suelo o
una rama cercana, deteniéndose el menor tiempo posible y cogiendo una sola
bellota lo más rápidamente posible. Los individuos más recelosos retiraban
bellotas de los comederos “al vuelo”, sin llegar a posarse en ellos. Otros
individuos nunca llegaron a vencer su miedo y, tras varios intentos infructuosos
de posarse en el comedero, desistieron de retirar bellotas. Esta perturbación
en el comportamiento natural de dispersión pudo provocar que infravaloráramos
el potencial de dispersión de la urraca debido a 1) la fracción de la población
que por miedo al comedero no dispersa bellotas desde el mismo; 2) la fracción
de la población que dispersa pocas bellotas desde los comederos por su
nerviosismo; y 3) todas las urracas no dispersan bellotas de los comederos
hasta que se han acabado las bellotas en el campo. Es decir, empiezan el
experimento con el almacén de bellotas “naturales” lleno, lo que pudo disminuir
la tasa de almacenamiento de las bellotas de los comederos incluso en las
urracas menos recelosas.
Aunque la cámara fue menos crítica
que el comedero, la posición de la cámara también influyó en el comportamiento
de la urraca. Cuando la cámara se fijó en el suelo (en la base de un tronco o
una piedra) perturbó menos el comportamiento que cuando se instaló elevada, en
el tronco o las ramas de la encina, a la altura del comedero o por encima. La
distancia mínima de las cámaras al comedero fue de 1.2 – 1.5 m.
Interacción abejero-velutina
Para analizar los cambios en el
comportamiento del abejero en presencia de las cámaras de fototrampeo
realizamos tres aproximaciones. En primer lugar, analizamos si la tasa de
reocupación de nidos de abejero de la temporada anterior se vio afectada por la
presencia de las cámaras en los nidos. Para ello, comparamos las tasas de
reocupación de los nidos en el periodo 2018-2020 (un 25%), donde los nidos no
tenían cámaras instaladas cuando llegaron los adultos en mayo, con las tasas de
reocupación en el periodo 2021-2022 (un 17.6%), donde los nidos tenían dos
cámaras instaladas (Browning Spec Ops y Spromise S378 4G). Aunque la tasa de
reocupación de nidos fue menor en el periodo 2021-2022, no hubo diferencias
significativas entre ambos periodos (c2
= 0.27, p = 0.68, n = 33 nidos, el p valor fue simulado a partir de 2000
iteraciones por el método de Montecarlo). La Chi-cuadrado utilizó los conteos
de nidos ocupados y no ocupados del periodo 2018-2020 para estimar las
frecuencias esperadas, y el conteo de nidos ocupados y no ocupados del periodo
2021-2022 para estimar las frecuencias observadas.
En segundo lugar, analizamos la respuesta de los abejeros a
la presencia de las cámaras instaladas en julio, cuando había pollos en el
nido. No aparecieron fotos donde los pollos miraran fijamente a la cámara el
mismo día o los días siguientes a su instalación, lo que sugiere que los pollos
ni siquiera se percataron de la presencia de las cámaras, lo que suele ocurrir
en las rapaces (García-Salgado et al. 2015). En el caso
de los adultos, en el 96.7% de los casos algún miembro de la pareja entró poco
después de instalar la cámara, observándose un retraso medio de 143 minutos
(rango 1 – 964 minutos). Sólo en uno de los nidos los adultos no entraron hasta
dos días después. Aunque las hembras entraron a los nidos antes que los machos,
las primeras con un retraso medio de 213.3 minutos (rango 1 - 964 minutos) y
los segundos de 402.6 minutos (rango 1 – 1589 minutos), no hubo diferencias
significativas en el test de Wilcoxon (W = 309, p = 0.35, n = 28 nidos). En el
86.2% de los casos, la hembra durmió con los pollos desde la primera o la
segunda noche tras la instalación de la cámara. Podríamos considerar ese
porcentaje (86.2%) como el porcentaje de parejas que se habituaron a la cámara
el mismo día o al día siguiente de su instalación. En un caso la hembra no
durmió hasta cinco días después de instalar la cámara en el nido. Finalmente,
en tres casos (10.3% de los casos) la hembra no durmió nunca en el nido una vez
instalada la cámara. En uno de los tres casos la hembra tuvo un comportamiento
diurno normal a partir de los cuatro días de la instalación. En los otros dos
casos la hembra no regresó al nido, pudiendo ser casos de deserción de la
hembra o que la hembra ya no estuviese presente cuando se instaló la cámara. En
ningún caso hemos detectado deserción del nido por los machos adultos.
En tercer lugar, comparamos casos donde instalamos una
cámara o dos cámaras el mismo día. El retraso en la entrada de los adultos al
nido fue similar (sin diferencias significativas en el test de Wilcoxon, W =
90.5, p = 0.76, n = 28 nidos), retraso medio de 132.4 minutos (rango 1 – 461
minutos) en los nidos con una cámara y de 155.1 minutos (rango 1 – 964 minutos)
en los nidos con dos cámaras. Los tiempos de habituación a las cámaras fueron
menores en los nidos donde instalamos una sola cámara, aunque las diferencias
no fueron significativas. Así, el 93.3% de las hembras durmieron en el nido la
primera o segunda noche tras instalar una cámara, mientras que el 78.6% de las
hembras lo hicieron en los nidos con dos cámaras (c2 = 1.33, p = 0.33, n = 29 nidos, el p valor fue simulado
a partir de 2000 iteraciones por el método de Montecarlo). La Chi-cuadrado utilizó
los conteos de hembras habituadas y no habituadas en los nidos con una cámara para
estimar las frecuencias esperadas, y el conteo de hembras habituadas y no
habituadas de los nidos con dos cámaras para estimar las frecuencias observadas.
Los tres casos donde la hembra no durmió posteriormente en el nido con los
pollos (incluidos los dos casos de posible deserción de la hembra) ocurrieron
en nidos donde instalamos dos cámaras.
Nuestros resultados muestran que la instalación de cámaras
de fototrampeo en los nidos de abejero altera el comportamiento de los adultos.
Sin embargo, en la mayoría de los casos (86.2%), los adultos se habitúan
rápidamente a la presencia de las cámaras, en el mismo día o al día siguiente
de su instalación. Ese porcentaje asciende al 93.3% si se instala una sola
cámara. Los mayores problemas aparecieron cuando instalamos dos cámaras
simultáneamente, lo que pudo influir en la tasa de deserción de la hembra adulta.
La deserción de las hembras de las rapaces es un fenómeno poco estudiado (Sonerud et al. 2014). Es más común en las
rapaces cuyos pollos consumen presas que no requieren la manipulación previa
del alimento por parte de la hembra (Eldegard et al. 2003). En ese caso, una vez
que los pollos pueden termorregular y dormir solos, la hembra puede desertar, y
el final de la crianza de los pollos recae únicamente en el macho. Este podría
ser el caso del abejero. Los pollos se alimentan principalmente de larvas y
pupas que no requieren manipulación previa por parte de la hembra porque los
pollos las consumen directamente de los panales que los adultos dejan en el
nido. Los vertebrados, que representan algo menos del 20% de la dieta de los
pollos de abejero, son de pequeño tamaño y edad juvenil y pueden ser fácilmente
manipulados por los pollos e incluso tragados directamente sin despedazar (Rebollo et al. 2023; Martín‐Ávila
et al. 2024). En estas circunstancias, una vez que los pollos son
capaces de termorregular, la instalación de cámaras puede favorecer un adelanto
de la deserción de la hembra adulta. Las hembras adultas de abejero inician la
migración postnupcial antes que los machos (Cento et al. 2021), lo cual puede tener que ver
con que las hembras de manera natural abandonan el cuidado de su descendencia
antes que los machos adultos. Nuestros resultados indican que las cámaras de
fototrampeo son muy valiosas para estudiar la dieta de las rapaces, pero cuando
se instalan en la proximidad de los nidos pueden alterar el comportamiento de
las aves adultas y perjudicar a las crías. En García-Salgado
et al. (2015) analizamos este tipo de problemas en las rapaces y
sugerimos soluciones para minimizar los impactos, entre otras, utilizar cámaras
de pequeño tamaño, poco conspicuas, instalar las cámaras cuando los pollos ya
tienen capacidad de termorregular por ellos mismos, y dar alimento
suplementario a los pollos en el momento de la instalación para protegerlos del
posible retraso en la entrada de los adultos. Los
resultados del presente artículo indican que hay que maximizar las precauciones
cuando se instala más de una cámara en el nido.
Problemas técnicos detectados en las cámaras de
fototrampeo
Interacción córvido-bellota
Los problemas técnicos detectados
pudieron solucionarse en un periodo muy corto, dada la accesibilidad de las
cámaras, situadas en el suelo o a poca altura en las encinas (Fig. 1), y la elevada
frecuencia de las revisiones: cada siete días en los estudios con arrendajos y
cada 1-3 días en los estudios con urracas. El agotamiento de las baterías, el
llenado de la tarjeta de memoria o los fallos de encuadre no fueron problemas
relevantes. Sin embargo, fue crítica la orientación de la cámara. La mejor
orientación fue hacia el norte, evitando el deslumbre del sol a primeras y
últimas horas del día. Otros problemas de visión, como la niebla o el objetivo
empañado por rocío a primeras horas de la mañana o últimas de la tarde, no se
pudieron solucionar, ocasionando una pequeña pérdida de información (problemas
en la identificación de especies, individuos o en el recuento de bellotas
removidas en los vídeos). La rapidez con que algunas urracas remueven las
bellotas del comedero, por su neofobia, exige disponer de cámaras con corto
tiempo de respuesta desde que se detecta el movimiento hasta que se inicia la
grabación del vídeo. La rapidez de las aves retirando las bellotas hizo que en
muchos casos la información que necesitábamos estuviese sólo en los primeros
fotogramas del vídeo, exigiendo vídeos con alta calidad para detectar la
remoción de las bellotas (1920 × 1080 píxeles y baja compresión de
almacenamiento o calidad “Ultra”). En relación con estos problemas, el modelo
Browning Dark Ops HD Elite funcionó razonablemente bien. Una característica
interesante de este modelo es que, teóricamente, mientras detecta movimiento no
interrumpe la grabación del vídeo (p. ej. durante el tiempo que el animal está
en pantalla eligiendo la bellota, hasta un máximo de dos minutos). De esta
manera, se evita que se pierda información si el animal agota el minuto
establecido previamente. Sin embargo, esta función no fue efectiva para
la urraca en la mayor parte de los casos, ya que es un animal pequeño y cubre
un porcentaje bajo de píxeles que no son suficientes para mantener activo el
sensor de movimiento y la cámara grabando. En el lado opuesto,
todos los modelos de cámaras produjeron un gran número de falsos positivos los
días de viento, grabándose vídeos sin presencia de córvidos debido al
movimiento de los árboles al fondo del encuadre. Esto puede llenar la memoria
de almacenamiento si las revisiones no son frecuentes, aumentando el tiempo
dedicado a la observación de los vídeos. Los nuevos programas que emplean
Inteligencia Artificial para filtrar imágenes podrían ayudar a reducir el
tiempo de revisión de los vídeos (Rodas-Trejo 2024).
Interacción abejero-velutina
El objetivo era que las
cámaras estuvieran activas de forma continua desde su instalación hasta la
salida de los volantones del nido, lo que evitaba escalar de nuevo a los nidos.
Con las Browning Spec Ops (33 cámaras) conseguimos este objetivo en el 78.8% de
los casos. Los problemas de funcionamiento de las cámaras se concentraron en
los dos primeros años (2018 y 2019, 10 cámaras), cuando sólo el 40% de las
cámaras permanecieron activas hasta la salida de los volantones, funcionando
una media de 20.3 días cada cámara. En el periodo posterior (2020-2022, 23
cámaras), el 95.6% de las cámaras funcionaron correctamente hasta la salida de
los volantones, estando operativas una media de 38.7 días. Los principales
fallos técnicos que interrumpieron el funcionamiento fueron el suministro
energético (cuatro casos, tres de ellos por
agotamiento de la batería externa y uno por entrada de agua en la batería
externa), llenado de la tarjeta de memoria (un caso), fallo en el soporte que
fijaba la cámara a la rama (un caso) y fallo interno desconocido de la cámara
(dos casos). Además, en cinco casos detectamos lapsus en el funcionamiento de
la cámara (periodos más o menos largos donde la cámara no hacía fotos a pesar
de la entrada de adultos al nido), lo que mermó su rendimiento, pero no
interrumpió el funcionamiento de la cámara. Estos lapsus estaban relacionados
principalmente con problemas en el suministro de energía y solían preceder a un
parón definitivo de la cámara por falta de ésta.
Las Spromise S378 4G (17 cámaras) funcionaron correctamente
hasta la salida de los volantones del nido en el 88.2% de los casos, estando
operativas una media de 37.1 días. Dos tipos de fallos interrumpieron su
funcionamiento: el suministro energético (dos casos) y la conexión de la
tarjeta SIM (dos casos). Sólo en un caso detectamos lapsus en el funcionamiento
de estas cámaras, lo que mermó su rendimiento, pero no interrumpió su
funcionamiento.
Además de los fallos técnicos descritos, cometimos errores
en la instalación de las cámaras Browning Spec Ops principalmente durante los
dos primeros años (2018 y 2019). El principal error fue la instalación de la
cámara demasiado cerca del nido, lo que provocó problemas de enfoque de las
fotos en cinco ocasiones. Además, esta cercanía provocó que el sensor de
movimiento se activara continuamente y la cámara hiciera fotos con demasiada
frecuencia, provocando fallos en el suministro energético y el llenado rápido
de la tarjeta de memoria, limitando el tiempo de funcionamiento. Otros dos
errores fueron el encuadre parcial del nido (siete casos) o la presencia de
obstáculos entre la cámara y el nido (tres casos), lo que redujo el campo de
visión de la cámara y la calidad de las fotos. Estos dos errores en ocasiones
fueron inevitables debido a la falta de lugares más adecuados para instalar la
cámara en las ramas situadas en las proximidades del nido (Fig.
4).
En resumen, podemos afirmar que la buena calidad actual de
las cámaras de fototrampeo, la reducción del tamaño haciéndolas menos
conspicuas y la reacción rápida ante los problemas (especialmente en el
suministro de energía) permiten rendimientos muy elevados en comparación con
los rendimientos de hace sólo una década (ver revisión en García-Salgado et al. 2015). Las cámaras
actuales tienen bajos consumos de energía y admiten tarjetas de memoria de alta
capacidad, lo que reduce los dos principales problemas que tenían los modelos más
antiguos. Los problemas se minimizan todavía más con las cámaras que envían las
fotos directamente al ordenador, como las Spromise S378 4G, permitiendo que los
investigadores reaccionen rápidamente ante los problemas.
Síntesis y conclusiones
Nuestros resultados muestran que las técnicas de fototrampeo
son muy útiles para el estudio de los servicios ecosistémicos suministrados por
la avifauna. Facilitan la identificación de la avifauna
que lleva a cabo el control de plagas y dispersión de semillas, así como la caracterización
y cuantificación de estos servicios ecosistémicos, a escala de población e
incluso de individuo. Los distintos modelos de cámaras permiten obtener
buenos resultados en condiciones ambientales contrastadas, es decir, flexibilidad
y adaptación a las circunstancias. Los investigadores deben identificar las
técnicas de fototrampeo que mejor se ajustan a los objetivos de su
investigación, a las especies diana y a las circunstancias de trabajo,
especialmente en aquellos casos en los que el acceso de los investigadores a
los sistemas de fototrampeo es difícil, complicando la detección y resolución
de los problemas.
El fototrampeo puede ser fácilmente combinado con otras
técnicas generando sinergias que permiten mejorar los resultados y profundizar
en la caracterización y cuantificación de los servicios que realizan las aves.
Las posibilidades son enormes y en este trabajo mostramos ejemplos exitosos de
combinación del fototrampeo con técnicas de marcaje de individuos y tecnologías
de seguimiento con emisores para el estudio de la dispersión de semillas
grandes y la regulación de plagas realizados por la avifauna. Esto ha
permitido, por ejemplo, obtener los primeros vídeos sobre la dispersión de
bellotas por urraca (Vídeos S3 a S14) o el ataque y consumo de
velutina por parte del abejero europeo (Vídeos S15 a S17). Actualmente
estamos extendiendo el estudio a otras especies de córvidos y de bellotas.
Sería interesante aplicar estas tecnologías con otros animales que almacenan
semillas y otras plantas en otros ecosistemas.
Debido a la naturaleza compleja de los procesos ecológicos
estudiados y de la tecnología empleada, el plan de trabajo debe ser flexible
(gestión adaptativa) para reaccionar frente a los imprevistos y solucionar los
problemas que surjan. Las técnicas de fototrampeo que envían las fotos o vídeos
directamente al ordenador permiten detectar en tiempo real los problemas e
identificar más rápidamente las soluciones. Hay que estar especialmente atento
a las interferencias de los equipos de fototrampeo en el comportamiento de la
especie diana. Aunque las cámaras y sus accesorios son cada vez más pequeños y
menos conspicuos, el fototrampeo es una técnica invasiva que puede modificar el
comportamiento de las aves, influir en los resultados y provocar efectos
negativos sobre las especies estudiadas, especialmente cuando se instalan en
las proximidades de los nidos. El plan de trabajo debe identificar rápidamente
estos impactos e iniciar la búsqueda de soluciones.
Contribución de los autores
Salvador Rebollo: Conceptualización, Adquisición de
financiación, Investigación, Metodología, Administración del proyecto,
Supervisión, Redacción – borrador original, Redacción – revisión y edición. Lorenzo
Pérez-Camacho: Conceptualización, Análisis de datos, Investigación,
Metodología, Supervisión, Visualización, Redacción – borrador original,
Redacción – revisión y edición. Loreto Martínez-Baroja: Conservación de datos, Análisis
de datos, Investigación, Metodología, Visualización, Redacción – revisión y edición.
Jorge A. Martín-Ávila: Conservación de datos, Análisis de datos, Investigación,
Metodología, Visualización, Redacción – revisión y edición. Luisa M.
Díaz-Aranda: Investigación, Metodología, Visualización, Redacción – revisión y
edición. Olivia Lorente-Casalini: Conservación de datos, Investigación,
Metodología, Redacción – revisión y edición. Noelia Morato: Investigación,
Redacción – revisión y edición. José M. Fernández-Pereira: Conservación de
datos, Investigación, Metodología, Redacción – revisión y edición. Jesús A.
Cuevas: Investigación, Metodología, Redacción – revisión y edición. José M.
Rey-Benayas: Adquisición de financiación, Administración del proyecto,
Redacción – revisión y edición. Pedro Villar-Salvador: Conceptualización,
Adquisición de financiación, Investigación, Metodología, Administración del
proyecto, Redacción – revisión y edición.
Disponibilidad de los datos
Todos los vídeos y sus leyendas están disponibles en: https://doi.org/10.6084/m9.figshare.28344395 (Rebollo et al.
2025).
Financiación, permisos requeridos, potenciales
conflictos de interés y agradecimientos
Agradecemos a Tomás González, Daniel Gómez, Pablo Quiles, Alberto
Pesqueira, Carlos Sobreira, José María Bello, Luis Ogando, Javier Orosa, María
Hernández, Marina López y Miguel A. Letón por sus contribuciones al trabajo de
campo y por proporcionar información; y al Ministerio de Defensa y al Real
Jardín Botánico Juan Carlos I por facilitar el acceso a las áreas de estudio.
Agradecemos a la Dirección Xeral de Conservación da Natureza de la Xunta de
Galicia y a la Comunidad Autónoma de Madrid por otorgarnos los permisos para
llevar a cabo el estudio. Este trabajo fue financiado por proyectos del CICYT
del Ministerio de Educación y Ciencia de España (CGL2007−60533/ BOS,
CGL2010−18312/BOS), el Ministerio de Economía y Competitividad de España
(CGL2014−53308−P) y el Ministerio de Ciencia e Innovación de España
(PID2019-106806GB-I00, PID2022-141762OB-I00), con fondos de la red REMEDINAL
(S−0505/AMB/0335, S2009 AMB−1783, S2013/MAE−2719, TE-CM S2018/EMT-4338) y un
proyecto de la Universidad de Alcalá (CCG2014/BIO−002). LMB fue financiada con
una Ayuda Postdoctoral Margarita Salas (Universidad de Alcalá), Ministerio de
Universidades de España y fondos NextGenerationEU. JAM-A fue apoyado por una
beca FPI (PRE2020-093652) del Ministerio de Ciencia, Innovación y Universidades
de España.
Los
autores/as declaran no tener ningún conflicto de intereses.
Referencias
Arca, M., Mougel, F., Guillemaud, T., Dupas, S., Rome,
Q., Perrard, A., Muller, F., et al. 2015. Reconstructing the invasion and the demographic
history of the yellow-legged hornet, Vespa velutina, in Europe. Biological
Invasions 17: 2357-2371. https://doi.org/10.1007/s10530-015-0880-9
Boesing, A.L., Nichols, E., Metzger, J.P. 2017. Effects of landscape
structure on avian-mediated insect pest control services: a review. Landscape
Ecology 32: 931-944. https://doi.org/10.1007/s10980-017-0503-1
Bommarco, R., Kleijn, D., Potts, S.G. 2013. Ecological intensification:
harnessing ecosystem services for food security. Trends in Ecology &
Evolution 28: 230-238. https://doi.org/10.1016/j.tree.2012.10.012
Budge, G.E., Hodgetts, J., Jones, E.P., Ostojá-Starzewski, J.C., Hall,
J., Tomkies, V., Semmence, N., et al. 2017. The invasion, provenance and
diversity of Vespa velutina Lepeletier (Hymenoptera: Vespidae) in Great
Britain. PLOS ONE 12: e0185172. https://doi.org/10.1371/journal.pone.0185172
Castro, J., Molina-Morales,
M., Leverkus, A.B., Martínez-Baroja, L., Pérez-Camacho, L., Villar-Salvador,
P., Rebollo, S., Rey-Benayas, J.M. 2017. Effective nut
dispersal by magpies (Pica pica L.) in a Mediterranean agroecosystem. Oecologia
184: 183-192. https://doi.org/10.1007/s00442-017-3848-x
Cento, M., Malpassuti, V.,
Dell’Omo, G., Agostini, N. 2021. Differential timing of autumn
migration between sex groups in adult European honey buzzards Pernis
apivorus. Avian Biology Research 14: 55-58. https://doi.org/10.1177/1758155921997364
Cramp, S.,
Simmons, K.E.L. eds. 1980. Handbook of the birds
of Europe, the Middle East and North Africa: the birds of the Western
Palearctic. Volume II. Hawks to Bustards. Oxford University Press, Oxford,
UK; New York, USA.
Eldegard, K., Selås, V., Sonerud, G.A., Steel, C., Rafoss, T. 2003. The
effect of parent sex on prey deliveries to fledgling Eurasian Sparrowhawks Accipiter
nisus. Ibis 145: 667-672.
https://doi.org/10.1046/j.1474-919X.2003.00229.x
Farwig, N.,
Berens, D.G. 2012. Imagine a world without seed
dispersers: A review of threats, consequences and future directions. Basic
and Applied Ecology 13: 109-115. https://doi.org/10.1016/j.baae.2012.02.006
Gamauf, A.,
Haring, E. 2004. Molecular phylogeny and
biogeography of Honey-buzzards (genera Pernis and Henicopernis). Journal
of Zoological Systematics and Evolutionary Research 42: 145-153. https://doi.org/10.1111/j.1439-0469.2004.00250.x
García, D., Zamora, R.,
Amico, G.C. 2010. Birds as suppliers of seed dispersal in
temperate ecosystems: conservation guidelines from real‐world
landscapes. Conservation Biology 24: 1070-1079. https://doi.org/10.1111/j.1523-1739.2009.01440.x
García, D., Miñarro, M.,
Martínez‐Sastre, R. 2021.
Enhancing ecosystem services in apple orchards: Nest boxes
increase pest control by insectivorous birds. Journal of Applied Ecology
58: 465-475. https://doi.org/10.1111/1365-2664.13823
García, D., Rumeu, B., Illera, J.C., Miñarro, M., Palomar, G.,
González-Varo, J.P. 2024. Common birds combine pest control and seed dispersal
in apple orchards through a hybrid interaction network. Agriculture,
Ecosystems & Environment 365: 108927. https://doi.org/10.1016/j.agee.2024.108927
García-Salgado, G.,
Rebollo, S., Pérez-Camacho, L., Martínez-Hesterkamp, S., Navarro, A.,
Fernández-Pereira, J.M. 2015. Evaluation of trail-cameras for
analyzing the diet of nesting raptors using the Northern Goshawk as a model. PLOS
ONE 10: e0127585. https://doi.org/10.1371/journal.pone.0127585
Gil-Pelegrín, E.,
Peguero-Pina, J.J., Sancho-Knapik, D. eds. 2017. Oaks
Physiological Ecology. Exploring the Functional Diversity of Genus Quercus L. Springer International Publishing, Cham, Switzerland.
Gómez, J.M., Schupp, E.W.,
Jordano, P. 2019. Synzoochory: the ecological and evolutionary
relevance of a dual interaction. Biological Reviews 94: 874-902. https://doi.org/10.1111/brv.12481
González-Varo, J.P.,
Albrecht, J., Arroyo, J.M., Bueno, R.S., Burgos, T., Escribano-Ávila, G.,
Farwig, N., et al. 2023. Frugivore-mediated seed dispersal in
fragmented landscapes: Compositional and functional turnover from forest to
matrix. Proceedings of the National Academy of Sciences 120:
e2302440120. https://doi.org/10.1073/pnas.2302440120
Hagemeijer,
W.J.M., Blair, M.J. eds. 1997. The EBCC atlas of
European breeding birds. Hauptw. Their distribution and abundance. 1. publ.
Poyser, London, UK.
Hougner, C., Colding, J., Söderqvist, T. 2006. Economic valuation of a seed
dispersal service in the Stockholm National Urban Park, Sweden. Ecological
Economics 59: 364-374.
https://doi.org/10.1016/j.ecolecon.2005.11.007
Landis, D.A. 2017. Designing agricultural landscapes for
biodiversity-based ecosystem services. Basic and Applied Ecology 18:
1-12. https://doi.org/10.1016/j.baae.2016.07.005
Liu, J., Dietz, T., Carpenter, S.R., Folke, C., Alberti, M., Redman,
C.L., Schneider, S.H., et al. 2007. Coupled human and natural systems. AMBIO:
A Journal of the Human Environment 36: 639-649. https://doi.org/10.1007/s13280-020-01488-5
Lundberg,
J., Moberg, F. 2003. Mobile link organisms and
ecosystem functioning: implications for ecosystem resilience and management. Ecosystems
6: 87-98. https://doi.org/10.1007/s10021-002-0150-4
Martín‐Ávila, J.A., Rebollo, S.,
Fernández‐Pereira, J.M.,
Díaz‐Aranda, L.M. 2024. The trophic strategy of the European honey‐buzzard Pernis
apivorus during breeding: extravagant specialization or ingenious solution?
Journal of Avian Biologye03221. https://doi.org/10.1111/jav.03221
Martín-Ávila, J.A., Díaz-Aranda, L.M., Fernández-Pereira, J.M., Rebollo, S.
2025. The European honey buzzard (Pernis apivorus) as an ally for the
control of the invasive yellow‐legged hornet (Vespa velutina nigrithorax). Pest
Management Science. https://doi.org/10.1002/ps.8622
Martínez-Baroja, L., Perez-Camacho,
L., Villar-Salvador, P., Rebollo, S., Quiles, P., Gomez-Sanchez, D.,
Molina-Morales, M., et al. 2019. Massive and effective acorn
dispersal into agroforestry systems by an overlooked vector, the Eurasian
magpie (Pica pica). Ecosphere 10: https://doi.org/10.1002/ecs2.2989
Martínez‐Baroja, L., Pérez‐Camacho, L., Villar‐Salvador, P., Rebollo, S.,
Leverkus, A.B., Pesendorfer, M.B., Molina‐Morales,
M., et al. 2021. Caching territoriality and site preferences
by a scatter‐hoarder drive the spatial pattern of seed dispersal and affect
seedling emergence. Journal of Ecology 109: 2342-2353. https://doi.org/10.1111/1365-2745.13642
Millennium Ecosystem
Assessment ed. 2005. Ecosystems and human
well-being: Synthesis. Island Press, Washington, DC, USA.
Molina-Morales, M., Albaladejo, G., Castro, J. 2019. Cache marking under field
conditions does not affect nut recovery rate by the Eurasian magpie Pica
pica, a scatter-hoarder corvid. Ardeola 66: 77-87. https://doi.org/10.13157/arla.66.1.2019.sc2
Molina-Morales, M.,
Castro, J., Albaladejo, G., Parejo, D. 2020. Precise cache
detection by olfaction in a scatter-hoarder bird. Animal Behaviour
167: 185-191. https://doi.org/10.1016/j.anbehav.2020.07.002
Molina‐Morales, M., Leverkus, A.B.,
Albaladejo‐Robles, G.,
Martínez‐Baroja, L.,
Pérez‐Camacho, L., Villar‐Salvador, P., Rebollo, S., et
al. 2023. Linking animal behaviour and tree recruitment:
caching decisions by a scatter hoarder corvid determine seed fate in a
Mediterranean agroforestry system. Journal of Ecology 111: 400-411. https://doi.org/10.1111/1365-2745.14004
Monteagudo, N., Benayas, J.M.R., Andivia, E., Rebollo, S. 2023. Avian
regulation of crop and forest pests, a meta‐analysis. Pest
Management Science 79: 2380-2389. https://doi.org/10.1002/ps.7421
Pérez-Camacho, L.,
Villar-Salvador, P., Cuevas, J.A., González-Sousa, T., Martínez-Baroja, L.
2023. Spatial decision-making in acorn dispersal by Eurasian
jays around the forest edge: Insights into oak forest regeneration mechanisms. Forest
Ecology and Management 545: 121291. https://doi.org/10.1016/j.foreco.2023.121291
Pesendorfer, M.B., Sillett, T.S., Koenig, W.D., Morrison, S.A. 2016.
Scatter-hoarding corvids as seed dispersers for oaks and pines: a review of a
widely distributed mutualism and its utility to habitat restoration. Condor
118: 215-237. https://doi.org/10.1650/CONDOR-15-125.1
Pons, J.,
Pausas, J.G. 2007. Acorn dispersal estimated by
radio-tracking. Oecologia 153: 903-911. https://doi.org/10.1007/s00442-007-0788-x
Rebollo, S., Rey-Benayas,
J.M., Villar-Salvador, P., Pérez-Camacho, L., Castro, J., Molina-Morales, M.,
Leverkus, A.B., et al. 2019. Services provided by birds
(high-mobile link species) in farmland and forest mosaics: forest regeneration
and plague regulation. Ecosistemas 28: 32-41. https://doi.org/10.7818/ECOS.1736
Rebollo, S., Díaz-Aranda,
L.M., Martín-Ávila, J.A., Hernández-García, M., López-Rodríguez, M.,
Monteagudo, N., Fernández-Pereira, J.M. 2023. Assessment of
the consumption of the exotic Asian Hornet Vespa velutina by the
European Honey Buzzard Pernis apivorus in southwestern Europe. Bird
Study 70: 136-150. https://doi.org/10.1080/00063657.2023.2244258
Rebollo, S., Pérez-Camacho, L.,
Martínez-Baroja, L., Martín-Ávila, J.A., Díaz-Aranda, L.M., Lorente-Casalini,
O., Morato, N., et al. 2025. Vídeos: Synergies between camera-trapping and
tracking technologies using transmitters for the study of ecosystem services
provided by birds / Sinergias entre el fototrampeo y tecnologías de seguimiento
con emisores para el estudio de servicios ecosistémicos suministrados por la
avifauna. Figshare. https://doi.org/10.6084/M9.figshare.28344395
Rodas-Trejo, J. 2024.
Camelot: Una herramienta intuitiva para el manejo y procesamiento de imágenes
de cámaras trampa utilizando inteligencia artificial. Ecosistemas 33:
2797. https://doi.org/10.7818/ECOS.2797
Rojas-Nossa,
S.V., Calviño-Cancela, M. 2020. The invasive hornet Vespa
velutina affects pollination of a wild plant through changes in abundance
and behaviour of floral visitors. Biological Invasions 22: 2609-2618. https://doi.org/10.1007/s10530-020-02275-9
Rome, Q., Muller, F.J., Touret‐Alby, A., Darrouzet, E.,
Perrard, A., Villemant, C. 2015. Caste differentiation and seasonal changes in Vespa
velutina (Hym.: Vespidae) colonies in its introduced range. Journal of
Applied Entomology 139: 771-782. https://doi.org/10.1111/jen.12210
Sonerud, G.A., Steen, R., Selås, V., Aanonsen, O.M., Aasen, G.-H.,
Fagerland, K.L., Fosså, A., et al. 2014. Evolution of parental roles in
provisioning birds: diet determines role asymmetry in raptors. Behavioral
Ecology 25: 762-772. https://doi.org/10.1093/beheco/aru053
UE1143/2014. Reglamento (UE) no
1143/2014 del Parlamento Europeo y del Consejo, de 22 de octubre de 2014, sobre
la prevención y la gestión de la introducción y propagación de especies
exóticas invasoras. Diario Oficial de la Unión Europea OJ L 317,
4.11.2014, p. 35–55. http://data.europa.eu/eli/reg/2014/1143/oj.
Whelan, C.J., Wenny, D.G.,
Marquis, R.J. 2008. Ecosystem services provided by birds. Annals
of the New York Academy of Sciences 1134: 25-60. https://doi.org/10.1196/annals.1439.003