Tradicionalmente, se ha utilizado el fototrampeo para
determinar abundancias de especies cuyos individuos pueden ser identificados
mediante características naturales (por ejemplo, características de pelaje) o
marcas artificiales (por ejemplo, marcas auriculares). Las identificaciones
permiten la construcción de matrices de presencia-ausencia, que pueden ser
analizadas con los modelos de captura-marcaje-recaptura (Karanth 1995; Heilbrun
et al. 2006). Si bien los sistemas
de cámaras se utilizan sobre todo para detectar la presencia de
carnívoros (Kucera et al. 1995; Naves
et al. 1996; Foresman y Pearson 1998)
ya que en la mayoría de los casos la identificación individual es muy
complicada y la identificación errónea puede producir sesgos significativos en
las estimaciones de población (Trolle et al. 2008; Yoshizaki
et al. 2009). En este sentido, en los últimos años se ha avanzado en
este aspecto (Steenweg et al. 2017), desarrollándose
diversos sistemas que permiten aproximarse a la estimación de abundancias
absolutas sin necesidad de reconocimiento individual (Rowcliffe et al. 2008; Moeller et al. 2018; Miles
et al. 2024). Por tanto, el fototrampeo puede utilizarse tanto para
estimar la densidad poblacional usando métodos basados en la captura-recaptura
como para derivar índices de abundancia relativa, en función de la cantidad de
capturas por unidad de esfuerzo.
En el presente trabajo se pretende evaluar la adecuación del
fototrampeo como método de estimación de abundancias en el tejón europeo
Meles meles, con énfasis en las poblaciones de baja densidad. Los objetivos
específicos fueron: 1) comparar la eficacia de cuatro atrayentes en la
detectabilidad del tejón; 2) evaluar la probabilidad de detección de tejones en
zonas de baja y alta densidad mediante técnicas de fototrampeo; 3) comparar las
diferencias en las abundancias relativas de tejón estimadas mediante fototrampeo
y conteo de letrinas; y 4) analizar cómo las estimaciones de abundancia
relativa de tejón con ambas metodologías varían en función de la densidad de
tejón.
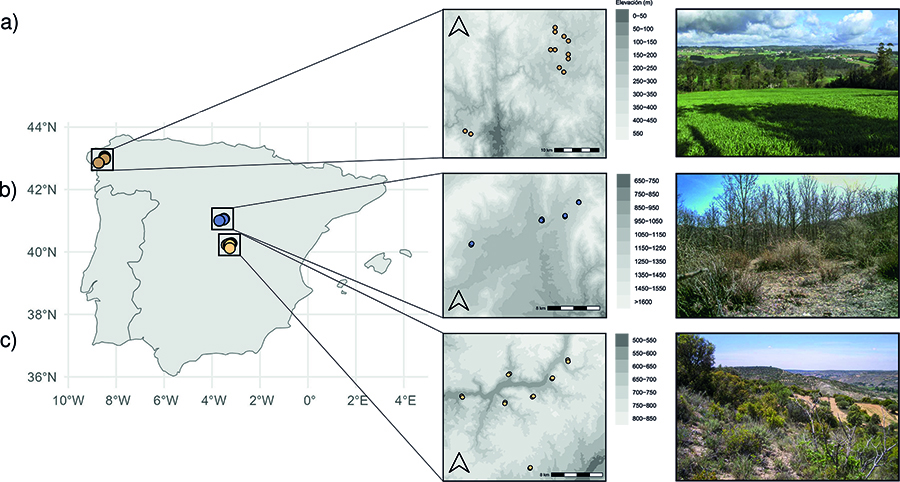
El estudio se llevó a cabo en
tres regiones distintas de la península ibérica, seleccionadas con objetivos
específicos y caracterizadas por distintas densidades de tejón (Fig. 1). Dos de ellas presentan altas densidades
poblacionales de tejón, mientras que la tercera se caracteriza por una densidad
baja. 1) Una región de estudio fue ubicada en la parte más oriental de la
Sierra de Guadarrama, la cual presenta una temperatura media anual de 10 ºC y
una precipitación media anual de 840 mm. Esta región presenta una elevada
densidad de tejones, con un registro medio de 3.24 letrinas por kilómetro (Escribano et al. 2009). En esta región, se seleccionaron
cuatro localidades, dos situadas a mayor altitud (1100-1150m): Montejo de la
Sierra (41° 3' 30” N, 3° 34' 44” O) y Prádena del Rincón (41° 2' 45" N, 3°
31' 40" O), donde predomina el roble melojo (Quercus pyrenaica) con
rodales de pino albar (Pinus sylvestris) y grandes extensiones de pastos
para uso de ganadería extensiva. Las otras dos localidades, Madarcos (41° 1'
52" N, 3° 34' 37 "O) y Villavieja del Lozoya (41° 0' 61" N, 3°
41' 49" O), están a menor altitud (≈ 1000m) y aunque predomina el roble
melojo también aparecen elementos más mediterráneos como encinas (Quercus
ilex); 2) Otra región de estudio se situó en el centro de la provincia de A
Coruña (43° 21' N, 8° 25' O).
Esta región está caracterizada por presentar una alta densidad de tejones,
pudiendo encontrar una media de 3.1 letrinas por kilómetro de transecto (Escribano et al. 2009). Esta región presenta
una temperatura media anual de 12.6 ºC y una precipitación media anual de 1886
mm. El paisaje está dominado por un mosaico agrosilvopastoral. Son
características las formaciones forestales de diversos tamaños en las que
predominan los cultivos de Eucalyptus globulus con algunas manchas
pequeñas de roble carballo (Quercus robur) que se entremezclan con
prados, cultivos y núcleos rurales donde abunda la ganadería extensiva de
vacuno. 3) Una última región de estudio se situó en la zona del sureste de
Madrid (40º 09'N, 3º 15'O). Presenta una temperatura media anual de 14.5 ºC y
una precipitación media anual de 380 mm. Esta región tiene muy baja densidad de
tejones y solo se conoce su presencia por citas puntuales (atropellos, huellas,
etc). En esta región predomina la vegetación xerofítica, caracterizada por
extensos espartales de Stipa tenacissima, intercalados con pequeños
rodales de bosque esclerófilo de encinas y coscojas (Quercus coccifera)
y bosques de Pinus halepensis. Estos parches están incluidos en una
matriz agrícola de uso intensivo y monoespecífico de plantaciones de olivo (Olea
europaea) y cultivos de secano, principalmente de cultivos de cereales.
Para comparar la eficacia de cuatro atrayentes en la
detectabilidad del tejón se llevó un experimento en la región de alta densidad
del norte de Madrid. Entre marzo y abril del 2012, se colocaron aleatoriamente,
y con una separación de entre 40 y 70 m, cuatro cámaras automáticas en una
superficie de 1x1 km2 en cada una de las cuatro localidades de la
región de alta densidad de tejones situadas en el norte de Madrid. En total se
colocaron 16 cámaras con sensor de movimiento ScoutGuard (12x), Bushnell
Trophy (2x) y Wildview Extreme 2 (2x). En cada una de las cuatro
cámaras se puso un atrayente diferente, aproximadamente a 1 m de distancia de
la cámara. Los atrayentes evaluados fueron seleccionados en base a su uso
tradicional en los estudios de carnívoros, y fueron los siguientes: (1) sardina
en lata, un atrayente generalista de carnívoros (Long
et al. 2008); (2) esencia de valeriana (Valeriana
officinalis), un atrayente que contiene ácido valérico presente en la orina
y las secreciones de las glándulas anales de algunos carnívoros (Saunders y Harris 2000; Schlexer 2008); (3) orina de lince ibérico Lynx
pardinus (obtenido de individuos en cautividad), que según Monterroso et
al. (2011)
promueve la curiosidad de la mayoría de los carnívoros al tratarse de un
competidor y posible depredador; y (4) agua de colonia comercial (Impacto),
ya que se ha observado que los carnívoros pueden sentirse atraídos por estos
olores (Moreira-Ramírez et al. 2011).
Para evaluar la probabilidad de detección de tejones en
zonas de baja y alta densidad, se realizó un estudio comparativo en las otras
dos regiones: la de baja densidad en el sureste de Madrid y la de alta densidad
en Galicia. En Madrid, se seleccionaron 7 localidades separadas al menos por 2
km entre ellas las cuales fueron seleccionadas en función de presencias
conocidas (atropellos, rastros, etc) de manera que, aunque fueran de muy baja
densidad, al menos se tuviera constancia de la presencia de la especie. El 18
de abril se colocaron dos cámaras en cada localidad (ScoutGuard (12x), Bushnell
Trophy (2x) de manera aleatoria), en una superficie de 1x1 km2 y
fueron georreferenciadas. En este caso, los atrayentes seleccionados fueron la
sardina y la colonia, de manera simultánea en las 14 cámaras y se mantuvieron
durante 35 días para maximizar la probabilidad de detección. Se repitió el
mismo proceso de revisión y renovación cada siete días que en el anterior
experimento. Por otro lado, el 20 de abril en la región de Galicia, se repitió
este mismo experimento sobre seis localidades. Concretamente se instalaron seis
cámaras Bushnell, dos Stealht Cam y cuatro cámaras Moultrie. La configuración
de las cámaras siguió el mismo protocolo descrito en el primer experimento,
manteniendo consistencia en los parámetros como el ángulo, altura, y tiempo de
funcionamiento para minimizar posibles sesgos metodológicos.
Para estimar las probabilidades de detección de tejones en
regiones con diferentes densidades se utilizaron modelos de ocupación basados
en métodos de máxima verosimilitud (MacKenzie et al. 2005), los cuales permiten
generar estimaciones de la probabilidad de ocupación considerando la detección
imperfecta. Para ello se utilizó el programa PRESENCE (Proteus Wildlife
Research Consultants, New Zealand; http://www.proteus.co.nz)
con el modelo para una temporada (MacKenzie et al. 2005). Este modelo utiliza
los muestreos temporales múltiples, para construir estimaciones apropiadas a
través de una serie de argumentos probabilísticos.
Para evaluar las diferencias significativas entre los
atrayentes en la detección de tejón, se ajustó un modelo lineal generalizado
inflado de ceros con errores de Poisson (Zero-Inflated Poisson, ZIP). Como
variable dependiente se utilizó el número de detecciones de tejones como
eventos independientes en las cámaras, mientras que el tipo de atrayente y la
localidad de la ubicación de la cámara fueron considerados como factores fijos
en el modelo. El efecto de los factores fijos se evaluó mediante una prueba de
razón de verosimilitud (LRT: Likelihood Ratio Test). Por tanto, se eliminaron
términos del modelo final si la comparación entre modelos con y sin un término
específico no resultó significativa según la prueba de LRT. Para examinar las
diferencias en la frecuencia de entradas de tejón atribuibles al tipo de
atrayente, se realizó un análisis post-hoc de Tukey. Este análisis permitió
evaluar cómo varía la frecuencia de detección en función del método de
detección dentro de cada región, considerando las interacciones entre estos
factores.
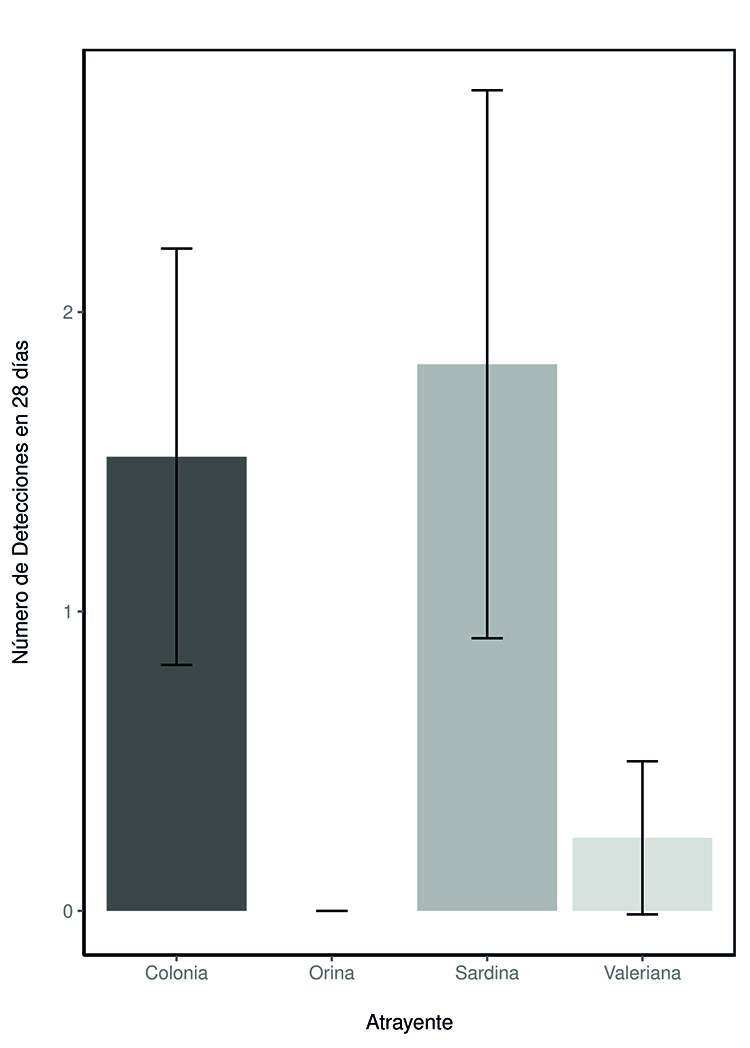
Figura 2. Número de detecciones de tejón (Meles
meles) durante los 28 días de experimento para los diferentes atrayentes:
colonia, orina de lince ibérico (Lynx pardinus), sardina en aceite y
esencia de valeriana (Valeriana officinalis). La figura muestra las
medias marginales junto con sus intervalos de confianza.
Figure
2. Number of European badger (Meles meles)
detections over the 28-day experiment for the different attractants: cologne,
Iberian lynx (Lynx pardinus) urine, canned sardines in oil, and valerian
(Valeriana officinalis) extract. The figure shows marginal means with
their confidence intervals.
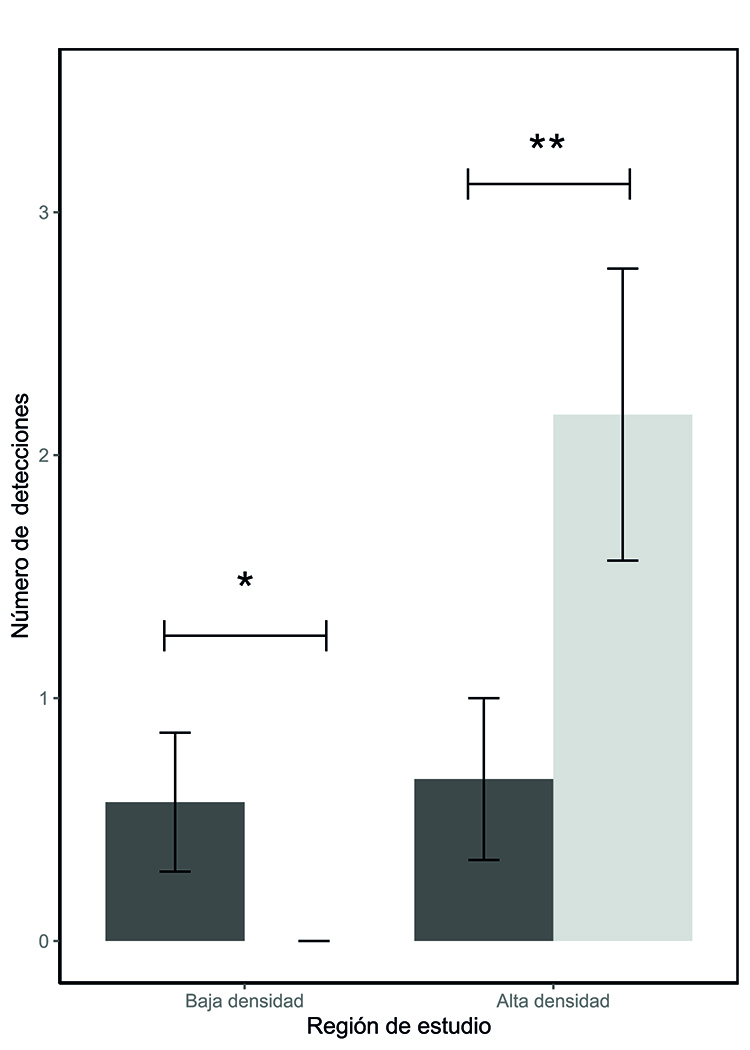
Figura 3.- Número de detecciones de tejón (Meles
meles) en función del método de muestreo (fototrampeo frente a conteo de
letrinas) en las regiones de baja (sureste de Madrid) y alta densidad
(Galicia). En gris oscuro están representadas el número de tejones detectados
con fototrampeo en 35 noches y en gris claro las detecciones mediante búsqueda
de letrinas en transectos de 2 km de longitud. La figura muestra las medias
marginales junto con sus intervalos de confianza.
Figure 3. Number of European badger (Meles meles) detections according
to the sampling method (camera trapping vs. latrine surveys) in regions of low
(southeastern Madrid) and high density (Galicia). Dark gray represents the
number of badgers detected through camera trapping over 35 nights, while light
gray represents detections obtained via latrine surveys along 2 km transects. The
figure shows marginal means with their confidence intervals.
Discusión
Nuestro estudio muestra que: 1) el fototrampeo combinado con
atrayentes fue un método adecuado para detectar tejones en nuestra región de
baja densidad, donde el recuento de letrinas fue ineficaz; 2) las estimaciones
de abundancia relativas obtenidas mediante fototrampeo no mostraron correlación
con los recuentos de letrinas; 3) la efectividad de un atrayente para una
especie particular depende del contexto (hábitat, clima, individuo), lo que
subraya la importancia de una cuidadosa selección y validación de los
atrayentes empleados.
El fototrampeo demostró ser una técnica eficaz para la
detección de tejones en áreas de baja densidad, donde el método indirecto de la
búsqueda de letrinas no resultó efectivo. Tradicionalmente, el recuento de
excrementos ha sido considerado el método más adecuado para la detección de
carnívoros (Barea-Azcón et al. 2007), y específicamente
de tejones (Tuyttens et al. 2001; Hutchings et al. 2002; Sadlier et al. 2004), debido a su eficiencia y
bajo costo. No obstante, diversos estudios han cuestionado su uso para la
evaluación de abundancias, especialmente en poblaciones de baja densidad (Harrington et al. 2010). El método basado
en el recuento de excrementos puede ser ineficaz por factores como el muestreo
no aleatorio, la incorrecta identificación de las heces o la variabilidad en la
detectabilidad debida a las diferentes tasas de descomposición (Barea-Azcón
et al. 2007). En ambientes mediterráneos, donde la densidad de tejones
suele ser baja, la tasa de descomposición microbiana de los excrementos es más
lenta debido a la menor humedad, lo que hace que las heces permanezcan más
tiempo en el campo y se acumulen en mayor cantidad (Sanchez et al. 2004). Esto podría aumentar la
probabilidad de detección, compensando parcialmente la menor abundancia de
animales (Cavallini 1994). No obstante, hay que tener en
cuenta que en medios más secos la actividad de los coprófagos es mayor y pueden
hacer desaparecer las heces muy rápido (Chiu et al. 2023). Estos aspectos deben ser
tenidos en cuenta cuando se comparan zonas con diferentes condiciones
climáticas. Para complicar más la detección, algunos estudios han observado que
los tejones no siempre sitúan las letrinas en los límites de sus territorios, y
el uso de las mismas es más esporádico e impredecible (Balestrieri et al. 2009). Nuestros
resultados, en línea con estos argumentos, sugieren que en zonas de baja
densidad podría ser necesario incrementar el esfuerzo de búsqueda. En
consecuencia, más estudios son necesarios para comprender mejor los patrones de
marcaje de los tejones y mejorar las estrategias de monitoreo en estos
entornos.
Aunque nuestros resultados de
detección en áreas de baja densidad podrían sugerir lo contrario, los
resultados indican que para usar el fototrampeo como método para estimar
la abundancia de tejones, es fundamental considerar múltiples factores que
influyen en su detectabilidad. Hemos observado que,
en poblaciones con densidades similares, como las de Galicia y el norte de
Madrid, se obtuvieron abundancias relativas muy dispares. De hecho, las
poblaciones de tejón del sureste de Madrid, a muy baja densidad, presentaron probabilidades
de detección en los modelos de ocupación más altas
que las registradas en Galicia, donde la densidad es mayor. Una fuente de
variación potencialmente importante en las probabilidades de detección es el
tamaño de la población (Kery 2002; Royle y Nichols 2003). Sin embargo, al
comparar poblaciones con densidades similares, encontramos grandes diferencias
en la detectabilidad. Esto sugiere que otros factores distintos a la densidad
poblacional están influyendo en estas variaciones. Por ejemplo, factores
ambientales como el clima o el hábitat, podrían estar afectando el número de
individuos activos y, por ende, la probabilidad de detección (O’Connell et al.
2011; Tanadini
y Schmidt 2011).
Además, es importante considerar que la actividad de
los tejones puede variar significativamente entre regiones. En Galicia, la
actividad de los tejones en marzo-abril podría ser menor que en el norte o el
sureste de Madrid debido a las diferencias climáticas y de latitud, lo que
podría reducir la probabilidad de detección. Esta menor actividad estacional
podría explicar por qué, a pesar de una mayor abundancia potencial, la
detectabilidad fue más baja en Galicia durante este periodo
Una posible explicación para las
diferencias en detectabilidad podría estar relacionada con las condiciones
tróficas de las poblaciones de tejones. En Galicia, las lombrices son una
fuente abundante de alimento, lo que podría hacer que los tejones no necesiten
recurrir a otras fuentes de alimento, y no se sientan atraídos por los
atrayentes utilizados en este estudio (Virgós
et al. 2004).
En cambio, en la región mediterránea, donde las lombrices son menos abundantes,
es plausible que los tejones dependan más de fuentes alternativas, como la
alimentación frugívora (Barea-Azcón
et al. 2010).
De este modo, las esencias vegetales presentes en las colonias comerciales
podrían estar favoreciendo la atracción de los tejones mediterráneos. No
obstante, se necesitan más investigaciones para comprender mejor los factores,
tanto comportamentales como ecológicos, que podrían estar influyendo en la
desvinculación entre la detectabilidad y la abundancia de la especie.
Numerosos investigadores parecen
coincidir en que el tejón es una especie que rehúye de los atrayentes y
que suele aparecer en baja frecuencia o incluso no aparece en estudios que
utilizan el trampeo fotográfico (Pardávila,
obs. propias; Barea-Azcón et al. 2007; Torre et al. 2009). La comparación entre los resultados que
obtuvieron Monterroso et al. (2011) y los que
se han obtenido en este estudio, muestra un claro ejemplo del grado de
variación en la atracción que presenta el tejón por los atrayentes,
especialmente en el caso de la orina de lince. La orina de lince es un buen
atrayente, puesto que el olor de un posible depredador promueve la curiosidad y
el marcaje de otros carnívoros (Monterroso et al.
2011). Sin embargo, mientras que Monterroso
et al. (2011) obtuvieron numerosas capturas, siendo el atrayente más
eficaz de los seis que evaluaron, tanto para tejón como para el resto de los
carnívoros ibéricos de la comunidad, en nuestro estudio la orina de lince no
facilitó la detección de tejones. Estas diferencias pueden ser debidas al
diferente origen de la orina, ya que Monterroso
et al. (2011) utilizaron orina de lince europeo (Lynx lynx),
mientras que en este estudio se utilizó orina de lince ibérico (Lynx
pardinus). No obstante, este factor no debería
representar un problema, dado que la orina de lince ibérico fue eficaz para el
resto de mesocarnívoros. Por otra parte, Burgos
et al. (2022)
encontraron que, aunque el lince ibérico condiciona la presencia de otros
mesocarnívoros, como la garduña, no parece alterar la presencia del tejón. Por
tanto, este resultado refuerza nuestro argumento sobre la importancia de
considerar las diferencias poblacionales al interpretar y diseñar estudios de
campo basados en fototrampeo. Además, pone de
manifiesto la dificultad de realizar comparaciones entre poblaciones debido a
las marcadas variaciones que pueden observarse incluso entre individuos de una
misma especie y a niveles de densidad similares.
Un
aspecto fundamental a considerar en los resultados obtenidos es la influencia
potencial de las características técnicas de las cámaras de fototrampeo.
Estudios recientes han mostrado que factores técnicos como la sensibilidad del
sensor, la velocidad de activación y el tipo de flash (infrarrojo o blanco)
pueden afectar a la detectabilidad (Palencia et al. 2022). Aunque en nuestro
estudio se emplearon diferentes marcas, se utilizaron cámaras con
especificaciones lo más similares posibles, lo que minimizó la variabilidad. Además,
las condiciones ambientales y climáticas propias de cada hábitat, como las
temperaturas extremas o la humedad, pudieron influir en el rendimiento de las
cámaras y, por ende, en la cantidad y calidad de los registros (Jacobs y Ausband 2018; McIntyre et al. 2020). Estas variaciones
técnicas y ambientales deben ser consideradas al interpretar los resultados, ya
que pueden tener un impacto significativo tanto en la detectabilidad como en
las estimaciones de abundancia relativa.
Conclusiones
Nuestros resultados destacan que el fototrampeo combinado
con atrayentes es una técnica eficaz para detectar tejones en áreas de baja
densidad poblacional. Este enfoque supera las limitaciones del conteo de
letrinas, el cual demuestra ser ineficaz cuando se aplica con la misma unidad
de esfuerzo y protocolo que en zonas de alta densidad. Sin embargo, aunque las
abundancias relativas fueron similares entre zonas con densidades
presumiblemente diferentes, estas variaciones podrían estar influenciadas por factores
adicionales, más allá de la densidad poblacional, como las condiciones
ambientales, las diferencias en el comportamiento de los tejones o incluso las
características específicas de las cámaras utilizadas. Nuestros hallazgos
subrayan la importancia de seleccionar y validar cuidadosamente los atrayentes
en función del contexto específico, ya que la eficacia de un atrayente puede
variar considerablemente entre regiones de estudio. Asimismo, destacan la
necesidad de ampliar las áreas de investigación y explorar el uso de atrayentes
alternativos para mejorar tanto la detección como las estimaciones de densidad.
En definitiva, nuestros resultados enfatizan la importancia de adaptar los
métodos de monitoreo a las características ecológicas y las densidades poblacionales
específicas de las especies objetivo. Esto es crucial para desarrollar
estimaciones robustas que permitan establecer relaciones confiables entre
abundancia y detectabilidad, optimizando así la eficacia de técnicas indirectas
como el fototrampeo en el monitoreo y la conservación de poblaciones
silvestres.
Contribución de los autores
Jesús López-Angulo: Conceptualización, Análisis formal,
Investigación, Metodología, Recursos, Redacción – borrador original,
Visualización. Xosé Pardavila-Rodríguez: Metodología, Recursos, Redacción –
revisión. Emilio Virgós: Conceptualización, Investigación, Metodología,
Recursos, Redacción – revisión, Supervisión, Validación.
Disponibilidad de datos
Los datos utilizados para este estudio están disponibles
públicamente a través de Zenodo, bajo el siguiente enlace: https://doi.org/10.5281/zenodo.14548813.
Financiación, permisos requeridos, potenciales
conflictos de interés y agradecimientos
Queremos agradecer a Vanesa Herranz por su valiosa ayuda en
la ejecución de los modelos de ocupación de una sola especie utilizando
PRESENCE, a Alberto Corral López por su apoyo en los conteos de letrinas, y a
los dos revisores anónimos por sus comentarios constructivos.
Los autores declaran que no tienen ningún
conflicto de intereses.
Referencias
Balestrieri, A.,
Remonti, L., Prigioni, C. 2009. Habitat selection in a
low-density badger meles meles population: A comparison of
radio-tracking and latrine surveys. Wildlife Biology 15: 442-448.
https://doi.org/10.2981/08-027
Barea-Azcón, J.M.,
Virgós, E., Ballesteros-Duperón, E., Moleón, M., Chirosa, M. 2007. Surveying carnivores at large spatial scales: A comparison of four
broad-applied methods. Biodiversity and Conservation 16: 1213-1230. https://doi.org/10.1007/s10531-006-9114-x
Barea-Azcón, J.M., Ballesteros-Duperón, E., Gil-Sánchez, J.M., Virgós, E. 2010.
Badger meles meles feeding ecology in dry mediterranean environments of the
southwest edge of its distribution range. Acta
Theriologica 55: 45-52. https://doi.org/10.4098/j.at.0001-7051.048.2008
Brown, J.A. 1993. Transmission of bovine tuberculosis (Mycobacterium bovis) from
badgers (Meles meles) to cattle. University of Bristol. Bristol, UK.
Bull, E.L., Holthausen, R.S., Bright, L.R. 1992. Comparison of three
techniques to monitor Marten. Wildlife Society Bulletin 20: 406-410.
Burgos, T., Fedriani, J.M., Escribano-Ávila, G., Seoane, J.,
Hernández-Hernández, J., Virgós, E. 2022. Predation risk can modify the
foraging behaviour of frugivorous carnivores: Implications of rewilding apex
predators for plant-animal mutualisms. Journal of Animal Ecology 91:
1024-1035. https://doi.org/10.1111/1365-2656.13682
Cavallini, P.
1994. Faeces count as an index of fox abundance. Acta
Theriologica 39: 417-424. https://doi.org/10.4098/AT.arch.94-49
Chiu, O., Tal, M., Sanmugam, A., Hesta, M., Gomez, D.E., Weese, J.S.,
Verbrugghe, A. 2023. The effects of ambient temperature exposure on feline
fecal metabolome. Frontiers in Veterinary Science 10: 1141881. https://doi.org/10.3389/fvets.2023.1141881
Clarke, G.P., White, P.C.L., Harris, S. 1998. Effects of roads on badger
Meles meles populations in south-west England. Biological Conservation
86: 117-124. https://doi.org/10.1016/S0006-3207(98)00018-4
Dekker,
J.J.A., Bekker, H.G.J. 2010. Badger (Meles meles)
road mortality in the Netherlands: the characteristics of victims and the
effects of mitigation measures. Lutra 53: 81-92.
Escribano, G., Virgós, E.,
Barja, I., Romero, C.L., Recio, M. 2009. Are populations in
the edge of their distribution range more stressed? A non-invasive field
evaluation of stress in the European badger (Meles meles) along an
environmental gradient in the Iberian Peninsula: Implications for global
change. Comparative Biochemistry and Physiology, Part A 2: S62. https://doi.org/10.1016/j.cbpa.2009.04.534
Ferreras, P., DÍaz-Ruiz,
F., Monterroso, P. 2018. Improving mesocarnivore detectability
with lures in camera-trapping studies. Wildlife Research 45: 505-517. https://doi.org/10.1071/WR18037
Foresman,
K.R., Pearson, D.E. 1998. Comparison of Proposed
survey procedures for detection of forest carnivores. The Journal of
Wildlife Management 62: 1217. https://doi.org/10.2307/3801985
Garrote, G., de Ayala, R.P., Pereira, P., Robles, F., Guzman, N., García,
F.J., Iglesias, M.C., et al. 2011. Estimation of the
Iberian lynx (Lynx pardinus) population in the Doñana area, SW Spain,
using capture-recapture analysis of camera-trapping data. European Journal
of Wildlife Research 57: 355-362. https://doi.org/10.1007/s10344-010-0440-7
Gese, E.M. 2001. Monitoring of terrestrial carnivore populations. Carnivore
Conservation 5: 372.
Griffiths,
H.I., Thomas, D.H. 1997. The conservation and
management of the European badger (Meles meles). Nature and
Environment Nº 90. Council of Europe Publishing, Strasbourg, France.
Harrington, L.A., Harrington, A.L., Hughes, J., Stirling, D., Macdonald, D.W.
2010. The accuracy of scat identification in distribution surveys: American
mink, Neovison vison, in the northern highlands of Scotland. European
Journal of Wildlife Research 56: 377-384. https://doi.org/10.1007/s10344-009-0328-6
Heilbrun, R.D., Silvy, N.J., Peterson, M.J., Tewes, M.E. 2006. Estimating
bobcat abundance using automatically triggered cameras. Wildlife Society
Bulletin 34: 69-73. https://doi.org/10.2193/0091-7648(2006)34[69:EBAUAT]2.0.CO;2
Hutchings, M.R., Service, K.M., Harris, S. 2002. Is population density
correlated with faecal and urine scent marking in European badgers (Meles
meles) in the UK? Mammalian Biology 67: 286-293. https://doi.org/10.1078/1616-5047-00043
Jackman, S., Tahk, A., Zeileis, A., Maimone, C., Fearon, J., Meers, Z.,
Jackman, M.S. 2015. Package 'pscl'. Political Science Computational
Laboratory. https://doi.org/10.32614/CRAN.package.pscl
Jacobs, C.,
Ausband, D. 2018. An evaluation of camera trap
performance - What are we missing and does deployment height matter? Remote
Sensing in Ecology and Conservation 4: 352-360. https://doi.org/10.1002/rse2.81
Karanth, K.U. 1995. Estimating tiger Panthera tigris populations from
camera-trap data using capture-recapture models. Biological conservation 71:
333-338. https://doi.org/10.1016/0006-3207(94)00057-W
Kays, R.W.,
Slauson, K.M. 2008. Remote cameras. In: Long, R.,
MacKay, P., Zielinski, W., Ray, J. (eds.), Noninvasive Survey Methods for
Carnivores: Methods and Analyses, pp. 110-140. Island Press, Washington,
USA.
Kery, M. 2002. Inferring the absence of a species: a case study of
snakes. The Journal of Wildlife Management 66(2), 330-338. https://doi.org/10.2307/3803165
Kucera, T.E., Soukkala, A.M., Zielinski, W.J. 1995. Photographic bait
stations. In: Ielinski, W. J., Kucera, T. E. (eds.), American Marten,
Fisher, Lynx, and Wolverine: Survey Methods for Their Detection, pp. 25-62.
US Department of Agriculture, Forest Service, Pacific Southwest Research
Station, Albany, CA, USA.
Kurek, P., Piechnik, Ł., Wiatrowska, B., Ważna, A., Nowakowski, K.,
Pardavila, X., Cichocki, J., et al. 2022. Badger Meles meles as ecosystem
engineer and its legal status in Europe. Animals 12: 1-19. https://doi.org/10.3390/ani12070898
Lara-Romero, C., Virgós,
E., Escribano-Ávila, G., Mangas, J.G., Barja, I., Pardavila, X. 2012. Habitat selection by European badgers in Mediterranean semi-arid
ecosystems. Journal of Arid Environments 76: 43-48. https://doi.org/10.1016/j.jaridenv.2011.08.004
Long, R.A., Zielinski, W.J., Long, R., MacKay, P., Zielinski, W., Ray,
J. 2008. Designing effective noninvasive carnivore surveys. Noninvasive
survey methods for carnivores 8-44.
Lopez-Angulo, J. 2024.
Datos de: Evaluación del fototrampeo como método de estimación de abundancias
en el tejón europeo Meles meles (Linnaeus, 1758). Zenodo. https://doi.org/10.5281/zenodo.14548813
MacKay, P., Zielinski, W.J.,
Long, R.A., Ray, J.C. 2008. Noninvasive research and
carnivore conservation. Island Press Washington,
DC, USA.
MacKenzie, D.I., Nichols, J.D., Sutton, N., Kawanishi, K., Bailey, L.L. 2005.
Improving inferences in population studies of rare species that are detected
imperfectly. Ecology 86: 1101-1113. https://doi.org/10.1890/04-1060
Martin-Garcia, S.,
Cortazar-Chinarro, M., Rodríguez-Recio, M., Jiménez, J., Höglund, J., Virgós,
E. 2023. Comparing minimum number of individuals and abundance
from non-invasive DNA sampling and camera trapping in the red fox (Vulpes
vulpes). Biodiversity and Conservation 32: 1977-1998. https://doi.org/10.1007/s10531-023-02586-y
McIntyre, T., Majelantle, T.L., Slip, D.J., Harcourt, R.G. 2020. Quantifying
imperfect camera-trap detection probabilities: Implications for density
modelling. Wildlife Research 47: 177-185. https://doi.org/10.1071/WR19040
Miles, V., Woodroffe, R., Donnelly,
C.A., Brotherton, P.N.M., Ham, C., Astley, K., Aurélio, J., et al. 2024.
Evaluating camera-based methods for estimating badger (Meles meles) density:
Implications for wildlife management. Ecological Solutions and Evidence 5:
1-15. https://doi.org/10.1002/2688-8319.12378
Moeller, A.K., Lukacs, P.M., Horne, J.S. 2018. Three novel methods to
estimate abundance of unmarked animals using remote cameras. Ecosphere
9: e02331. https://doi.org/10.1002/ecs2.2331
Monterroso, P., Alves,
P.Ć., Ferreras, P. 2011. Evaluation of attractants for
non-invasive studies of Iberian carnivore communities. Wildlife Research
38: 446-454. https://doi.org/10.1071/WR11060
Moreira-Ramírez, J., McNab, R., Garcia-Anleu, R., Méndez, M., Barnes, M.,
Ponce-Santizo, G., Córdova, M. 2011. Densidad de Jaguares dentro de
la Concesión Comunitaria de Carmelita y de la Asociación Forestal Integral San
Andrés Petén, Guatemala. Wildlife Conservation,
Society-Jaguar Conservation Program, Guatemala.
Naves, J., Fernández, A., Gaona, J.F., Nores, C. 1996. Uso de
cámaras automáticas para la recogida de información faunística. Doñana, Acta
Vertebrata 23: 189-199.
Neal, E.G., Cheeseman, C.L.
1996. Badgers. Poyser Natural History, London, UK.
Negroes, N., Sollmann, R.,
Fonseca, C., Jacomo, A.T.A., Revilla, E., Silveira, L. 2012. One
or two cameras per station? Monitoring jaguars and other mammals in the Amazon.
Ecological research 27: 639-648. https://doi.org/10.1007/s11284-012-0938-4
Newton, A.C., Johnson, S.N., Gregory, P.J. 2011. Implications of climate
change for diseases, crop yields and food security. Euphytica 179: 3-18.
https://doi.org/10.1007/s10681-011-0359-4
O'Connell, A.F., Nichols, J.D., Karanth, K.U. 2011. Camera traps in animal
ecology: methods and analyses. Springer. https://doi.org/10.1007/978-4-431-99495-4
Palencia, P., Vicente, J.,
Soriguer, R.C., Acevedo, P. 2022. Towards a best-practices
guide for camera trapping: assessing differences among camera trap models and
settings under field conditions. Journal of Zoology 316: 197-208. https://doi.org/10.1111/jzo.12945
Roper, T.J. 2010. Badger. Harpercollins Publishers, London, UK.
Rowcliffe, J.M., Field, J., Turvey, S.T., Carbone, C. 2008. Estimating animal
density using camera traps without the need for individual recognition. Journal
of Applied Ecology 45: 1228-1236. https://doi.org/10.1111/j.1365-2664.2008.01473.x
Royle, J.A.,
Nichols, J.D. 2003. Estimating abundance from
repeated presence-absence data or point counts. Ecology 84: 777-790. https://doi.org/10.1890/0012-9658(2003)084[0777:EAFRPA]2.0.CO;2
Sadlier, L.M.J., Webbon, C.C., Baker, P.J., Harris, S. 2004. Methods of
monitoring red foxes Vulpes vulpes and badgers Meles meles: Are field signs the
answer? Mammal Review 34: 75-98. https://doi.org/10.1046/j.0305-1838.2003.00029.x
Sanchez, D.M., Krausman, P.R., Livingston, T.R., Gipson, P.S. 2004.
Persistence of carnivore scat in the Sonoran Desert. Wildlife Society
Bulletin 32: 366-372. https://doi.org/10.2193/0091-7648(2004)32[366:POCSIT]2.0.CO;2
Sargeant, G.A.., Johnson, H.D., Berg, W.E. 1998. Interpreting carnivore
scent-station surveys. The Journal of Wildlife Management 62: 1235-1245.
https://doi.org/10.2307/3801987
Saunders,
G., Harris, S. 2000. Evaluation of attractants and
bait preferences of captive red foxes (Vulpes vulpes). Wildlife
Research 27: 237-243. https://doi.org/10.1071/WR99052
Schlexer, F.V.
2008. Attracting animals to detection devices.
Island Press Washington DC, USA.
Schley, L., Schaul, M., Roper, T.J. 2004. Distribution and population
density of badgers Meles meles in Luxembourg. Mammal Review 34:
233-240. https://doi.org/10.1111/j.1365-2907.2004.00040.x
Steenweg, R., Hebblewhite, M., Kays, R., Ahumada, J., Fisher, J.T., Burton,
C., Townsend, S.E., et al. 2017. Scaling-up camera traps: monitoring the
planet's biodiversity with networks of remote sensors. Frontiers in Ecology
and the Environment 15: 26-34. https://doi.org/10.1002/fee.1448
Tanadini,
L.G., Schmidt, B.R. 2011. Population size
influences amphibian detection probability: Implications for biodiversity
monitoring programs. PLoS ONE 6: e28244. https://doi.org/10.1371/journal.pone.0028244
Torre, I., Ribas, A.,
Arrizabalaga, A. 2009. Estudio de la comunidad de carnívoros del PN Del
Montseny (Catalunya) mediante trampeo fotográfico. Galemys 21: 165-180. https://doi.org/10.7325/Galemys.2009.NE.A13
Trolle, M., Noss, A.J.,
Cordeiro, J.L.P., Oliveira, L.F.B. 2008. Brazilian tapir
density in the Pantanal: A comparison of systematic camera‐trapping
and line‐transect surveys. Biotropica 40: 211-217. https://doi.org/10.1111/j.1744-7429.2007.00350.x
Tuyttens, F.A.M., Macdonald, D.W., Swait, E., Cheeseman, C.L. 1999.
Estimating population size of Eurasian badgers (Meles meles) using
mark-recapture and mark-resight data. Journal of Mammalogy 80: 950-960. https://doi.org/10.2307/1383265
Tuyttens, F.A.M., Long, B., Fawcett, T., Skinner, A., Brown, J.A.,
Cheeseman, C.L., Roddam, A.W., et al. 2001. Estimating group size and
population density of Eurasian badgers Meles meles by quantifying
latrine use. Journal of Applied Ecology 38: 1114-1121. https://doi.org/10.1046/j.1365-2664.2001.00665.x
Virgós, E., Mangas, J.G., Blanco-Aguiar, J.A., Garrote, G., Almagro, N.,
Viso, R.P. 2004. Food habits of European badgers (Meles meles) along an
altitudinal gradient of Mediterranean environments: a field test of the
earthworm specialization hypothesis. Canadian Journal of Zoology 82:
41-51. https://doi.org/10.1139/z03-205
Virgós, E., Revilla, E., Mangas, J.G., Domingo-Roura, X. 2005. Ecología
y conservación del tejón en ecosistemas mediterráneos. Sociedad Española
para la Conservación y Estudio de los Mamíferos (SECEM), Málaga, España.
Williams, B.K., Nichols, J.D., Conroy, M.J. 2002. Analysis and management
of animal populations. Academic press.
Wilson, G., Harris, S., McLaren, G. 1997. Changes in the British badger
population, 1988 to 1997. People's Trust for Endangered Species London, UK.
Woodroffe,
R., Redpath, S.M. 2015. When the hunter becomes the
hunted. Science 348: 1312-1314. https://doi.org/10.1126/science.aaa8465
Yoshizaki, J., Pollock, K.H., Brownie, C., Webster, R.A. 2009. Modeling
misidentification errors in capture-recapture studies using photographic
identification of evolving marks. Ecology 90: 3-9. https://doi.org/10.1890/08-0304.1