Introducción
El tapir andino (Tapirus pinchaque
Ruolin, 1829) es considerado como una especie clave en los páramos andinos por
su dieta herbívora y frugívora (Acosta
et al. 1996; Lizcano y Cavelier 2004a; Bermúdez
y Reyes Puig 2011). Su
distribución se restringe a los bosques montanos y páramos de Colombia (Acosta
et al. 1996) Ecuador
(Downer 1996) y norte de Perú (Padilla et al. 2010), en un rango altitudinal que va de los 1200 a los 4500 m (Tirira
et al. 2019; Castellanos et al. 2020). La especie se considera extinta en
Venezuela donde no hay registros desde hace más de 40 años (Acosta
et al. 1996; Downer 1996).
Aunque la información disponible sobre su situación actual es escasa, la dramática destrucción y degradación de sus hábitats debido a
la colonización, actividades extractivas, presión por caza y comercio ilegal de
sus partes han colocado al tapir andino como uno de los mamíferos más
amenazados de Sudamérica, considerado En Peligro Crítico en Ecuador (Urgilés-Verdugo
et al. 2021). Se
trata de la especie más amenazada del género Tapirus (Tirira
et al. 2019) y con menor densidad poblacional de las tres especies de tapires
del Neotrópico (Lizcano
y Cavelier 2000b).
La gran diversidad biológica,
ecosistémica y cultural de los bosques andinos, contrasta con su extremada
vulnerabilidad a los cambios en el uso del suelo y el clima (de la Torre
et al. 2006; Mena Vásconez y Hofstede
2006; Quichimbo et al. 2012; Tejedor Garavito et al.
2012). Su historia de uso
data de hace más de 3000 años para actividades agropecuarias (Hofstede
et al. 2002; de la Torre et al. 2006). En Ecuador, la región andina es la más
densamente poblada (Quichimbo
et al. 2012) con
menos del 3% del total de páramo ecuatoriano en buen
estado de conservación (Hofstede
et al. 2002; Quichimbo et al. 2012). Los remanentes bien conservados de
páramo y bosques montanos se encuentran en la vertiente oriental de la
cordillera (de la Torre
et al. 2006). El
resto del hábitat disponible por encima de los 3000 m s.n.m. en Ecuador, unos
8000 km2, han sido transformados por intervención humana (Hofstede
et al. 2002). Esta
pérdida de hábitat en Ecuador, representa una tasa de deforestación del 2%
anual entre 2005 y 2010, la más alta de todos los países andinos (Tejedor
Garavito et al. 2012). Estimaciones más actualizadas, contrastan en la cantidad
porcentual de pérdida de bosques montanos y páramos en el área de distribución
del tapir andino en Ecuador, por cambio en el uso del suelo y cambio climático,
del 2% (Mestanza-Ramón
et al. 2021) al 17% (Ortega-Andrade et al. 2015).
Los datos de densidad poblacional del
tapir andino son escasos y relativamente antiguos, tanto en Ecuador (Downer 1996; Castellanos
2013) como en Colombia (Acosta
et al. 1996; Lizcano y Cavelier 2000b). Los datos de radio collares obtenidos
en páramos de Ecuador dieron densidades que variaban entre 0.17 y 0.29
individuos/km2 (Downer 1996; Castellanos
2013). En el caso de Colombia, la información procedía del
análisis de huellas en páramos y las estimaciones de densidad fueron parecidas a
las registradas en Ecuador, de 0.18 a 0.25 individuos/km2 (Acosta
et al. 1996; Lizcano y Cavelier 2000b). Si bien la metodología de radio
collares GPS es robusta (O’Connell
et al. 2011),
implica dificultades logísticas, no está exenta de dificultades éticas por las
técnicas de captura y manejo de individuos (Downer 1996; Castellanos
2013), y además es
costosa (O’Connell
et al. 2011).
Las estimaciones generadas por registros
indirectos son pocos robustas, implican tiempo de campo y personal capacitado para
la toma de datos (Acosta
et al. 1996; Lizcano y Cavelier 2000b). En este sentido, las técnicas de
fototrampeo se han mostrado enormemente eficaces para obtener datos
informativos (Rovero
y Marshall 2009) y sin
las limitaciones de las técnicas de radio collares y de radios tracking (Efford 2004). Las estimaciones disponibles para el
tapir andino se llevaron a cabo en un solo tipo de hábitat, páramo o bosque
montano (Acosta et al. 1996; Downer 1996; Lizcano
y Cavelier 2000a; Castellanos 2013), lo cual no permite saber si existen diferencias de densidad de
tapir andino entre sus principales hábitats. Downer (1996) registró el uso
diferenciado de hábitat del tapir andino en el Parque Nacional Sangay, Ecuador,
en un 80% para los bosques montanos, mientras que el páramo fue utilizado en el
20% restante. Sin embargo, existen discrepancias sobre estos datos, dado que
otro autore sugiere que el tapir andino usa preferentemente el páramo (Castellanos 2013).
Además de un uso de hábitat diferenciado,
existen otros factores clave para explicar la densidad del tapir andino, como
son la interacción negativa con depredadores, como el puma (Puma concolor)
y el oso andino (Tremarctos ornatus) (Downer 1996), la bien conocida relación inversa
entre el tamaño promedio de los individuos con su densidad (Damuth 1981) y la existencia de perturbaciones
antrópicas (Acosta
et al. 1996). La
detectabilidad del tapir varía no solo con la abundancia, sino también con su
actividad que está asociada con el hábitat, el cual es usado también de forma
diferencial por los depredadores, pero también por animales exóticos
introducidos, como los perros ferales (Acosta
et al. 1996; Lizcano y Cavelier 2000b, b; Lizcano
et al. 2002; Zapata-Ríos y Branch 2016; Mena et al.
2020). Determinar si existen diferencias significativas
en la densidad poblacional del tapir andino entre sus dos principales hábitats es
algo prioritario para la planificación de estrategias de conservación de una
especie clave en estos ecosistemas amenazados. Para ello, es necesario utilizar
variables relacionadas con los condicionantes naturales, pero también con otras
que puedan indicar la existencia de perturbaciones antrópicas, como se ha hecho
para modelizar la distribución, estado de conservación y ocupación de otros grandes
ungulados y carnívoros (Ortega-Andrade et al. 2015; Zapata-Ríos y Branch 2018; Mena et al.
2020; Mestanza-Ramón et al. 2021). El objetivo de nuestro trabajo es estimar la densidad
poblacional del tapir andino en el noreste de los Andes de Ecuador con énfasis en
la variación y preferencia entre sus dos hábitats principales: bosque montano y
páramo; así como detectar aquellos factores ambientales y relacionados con las
perturbaciones antrópicas que afectan su ocupación (y) y
detectabilidad (p). Se hipotetiza que la densidad y ocupación del tapir
andino es mayor en el bosque montano que en páramo dado que la productividad y
la oferta de recursos nutricionales es mayor en este último hábitat (Downer 2001; Tejedor
Garavito et al. 2012).
Material y métodos
Área de estudio
El estudio se realizó en cuatro
localidades de la vertiente nororiental de los Andes ecuatorianos: Monte-Olivo,
Oyacachi y Río Cedro (ubicadas en el Parque Nacional Cayambe-Coca, PNCC), y
Cuyuja (en la Reserva Ecológica Antisana, REA) (Fig. 1). Monte-Olivo y Oyacachi
son páramos (ecosistema de herbazal de páramo), mientras Cuyuja y Río Cedro son
bosques montanos (ecosistema de Bosque siempreverde montano alto del Norte de
la Cordillera Oriental de los Andes).
El ecosistema de páramo es heterogéneo e
incluye planicies de alta montaña (páramo herbáceo), páramo de rosetal
(almohadillas), páramo arbustivo y páramo de frailejones (Baquero
et al. 2004; Tejedor Garavito et al.
2012; Ministerio del Ambiente
del Ecuador 2013). De
manera similar, los bosques montanos presentan una notable heterogeneidad y
están conformados por bosque de neblina, montano y montano alto (Baquero
et al. 2004; Tejedor Garavito et al.
2012; Ministerio del Ambiente
del Ecuador 2013). En
cada localidad, se seleccionó un área de 10 x 10 km (100 km²) representativa de
hábitats de páramo y bosque montano, sin evidencias aparentes de perturbación
antrópica. Las parcelas estaban separadas entre 5 a
10 km para garantizar independencia espacial. El tamaño de las parcelas
respondió a características ecológicas y de comportamiento del tapir andino,
como su ámbito hogareño que fue estimado entre 2.5 a 3.5 km2 (Lizcano
y Cavelier 2004b). Las
coordenadas geográficas del centroide de cada parcela y su altitud promedio fueron,
para ecosistema de páramo: Monte Olivo (00°20’ N, 77°49’W,
3563 msnm) y Oyacachi (00°13’ S, 78°02’ W, 3555 msnm); para
ecosistema de bosque montano: Cuyuja (00°28’ S, 78°03’W, 2966 msnm) y
Río Cedro (00°15’ S, 77°54’ W, 2758 msnm). La temperatura media mensual en el páramo es de 6 °C y en
bosque montano de 9.5 °C. La precipitación media mensual durante los meses
de muestreo fue 117 mm en páramo y 150.6 mm en bosque montano (Ministerio del Ambiente del Ecuador 2013). Las localidades de Oyacachi y Cuyuja
presentan asentamientos humanos a una distancia aproximada de 10 km en línea
recta del borde de la parcela.
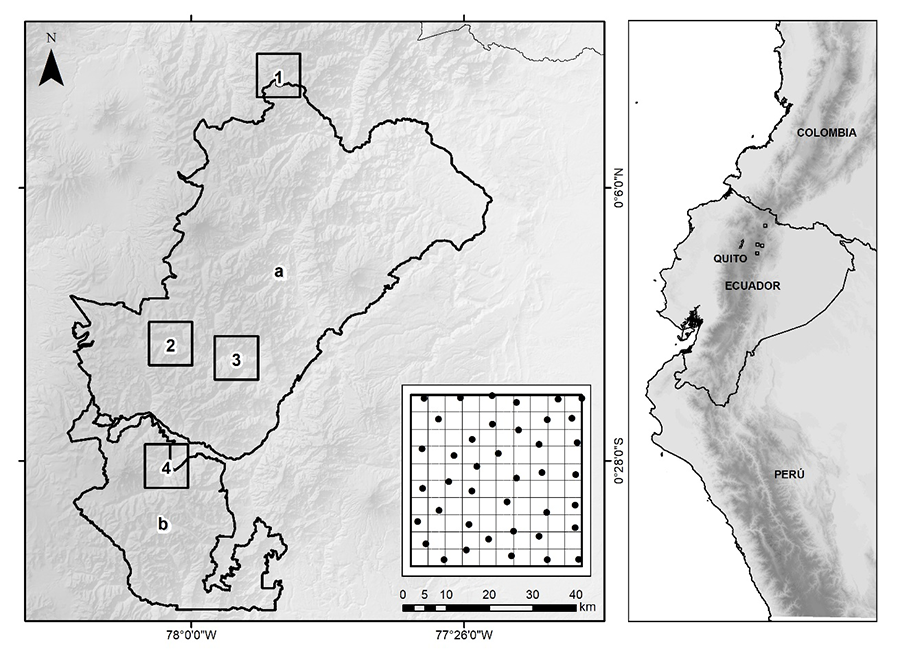
Figura 1. Mapa de área de
estudio. Se incluye las dos regiones de estudio: a. Parque Nacional Cayambe
Coca, b. Reserva Ecológica Antisana. Los cuadrantes negros indican las
localidades de muestreo: 1. Monte Olivo, 2. Oyacachi, 3. Río Cedro, 4. Cuyuja.
Los puntos negros de cada malla corresponden a las ubicaciones concretas de las
cámaras trampa.
Figure
1. Map of the study area. The two study areas are
included: a. Cayambe Coca National Park, b. Antisana Ecological Reserve. The
black quadrants indicate the sampling localities: 1. Monte Olivo, 2. Oyacachi,
3. Río Cedro, 4. Cuyuja. In the grid, the black dots correspond to the specific
locations of the camera traps.
Toma de datos
Mediante fototrampeo evaluamos la
abundancia del tapir andino entre septiembre de 2011 y febrero de 2013 (18
meses). En cada parcela de 10 x 10 km establecimos una malla de 100 celdas de 1
x 1 km. Seleccionamos al azar 40 de estas celdas y colocamos una cámara
trampa en cada una de ellas (Bushnell Trophy Cam,
119456 C) separadas por al menos 2.5 km de distancia
entre ellas. Esta distancia mínima se basó en las
estimaciones del ámbito hogareño del tapir andino (Lizcano
y Cavelier 2004b). Las
cámaras permanecieron en cada sitio, en promedio, 70.8
días SD ± 8.3 y se mantuvieron activas durante 24 horas. Se tomaron tres fotografías
por evento de captura a intervalos de tres minutos.
Para minimizar la probabilidad de sobrestima por recapturas incidentales se
consideraron eventos independientes de captura fotográfica las fotografías que
cumplían los siguientes criterios: a. fotografías consecutivas de distintos
individuos de la misma especie, b. fotografías no consecutivas de individuos de
la misma especie (O’Brien
et al. 2003) y c.
fotografías consecutivas de individuos de la misma especie tomadas con al menos
60 minutos de diferencia (O’Brien
et al. 2003; Jiménez et al. 2010; Carbajal-Borges et al.
2014). Este planteamiento
es más conservador que otras propuestas metodológicas, como O’Brien et al. (2003), quienes consideraron exclusivamente fotografías consecutivas de
individuos de la misma especie tomadas con al menos 30 minutos de diferencia.
Los factores relacionados con la
ocupación (y) y la densidad (p) del tapir andino fueron estimados mediante modelos de ocupación (MacKenzie
y Bailey 2004; Mackenzie y Royle 2005), con variables de hábitat y de incidencia
de perturbación antrópica (Tabla
1). Las variables fueron evaluadas a dos
escalas, (a) local (estación de fototrampeo), que comprende una distancia
radial de 20 m alrededor de cada cámara trampa y (b) de paisaje entendida, como
el área total de cada parcela de muestreo de 100 km2 (Tabla 1).
A escala de paisaje estimamos la distancia más corta a zonas pobladas, fuentes
de agua y vías como subrogados de perturbación antrópica (Tabla 1).
Las variables de hábitat se evaluaron a la escala local, utilizando datos de
presencia de carnívoros obtenidos mediante las mismas cámaras trampa. Para el
análisis, se consideraron únicamente eventos de captura independientes. Los
carnívoros fueron incluidos en los modelos como potenciales depredadores (Downer 1996; Rodriguez
et al. 2014; Pisso-Florez et al. 2021). Las medidas de la temperatura ambiente
se obtuvieron de las cámaras trampa. La elevación se midió en campo. La
precipitación media mensual y pendiente se obtuvieron del mapa de ecosistemas
del ecuador (Ministerio del Ambiente del Ecuador 2013). La pendiente se clasificó en tres
categorías según su porcentaje de inclinación: ligera = 1-8% (a); moderada = 9-40%
(b); fuerte = +40% (c). Imágenes espectrales LANSAT-8 sirvieron para estimar
el índice de vegetación de diferencia normalizada (NDVI) como un estimador de
la producción primaria en cada localidad de fototrampeo con un tamaño de píxel
de 30 m. Las variables altamente correlacionadas (r > 0.5) se omitieron en la
elaboración de nuestros modelos.
Tabla 1. Variables
de hábitat y perturbación antrópica medidas en los Andes ecuatorianos para
estimar la ocupación (y) y
detectabilidad (p) del tapir andino. Se incluye el rango de valores de las
variables, escala en la que cada variable fue medida y fuente de los datos. El asterisco muestra las variables usadas en los modelos definitivos.
Table
1. Habitat and
anthropogenic disturbance variables measured in the Ecuadorian Andes to model
the Andean tapir occupancy (y)) and detectability (p). We also presented the range of variable
values, scale at which each variable was measured and source of data. The
asterisk shows the variables used in the final models.
Análisis de datos
Estimamos la densidad del tapir andino
mediante el Modelo de Encuentros Aleatorios (MEA) (Rowcliffe et al. 2008). El
MEA no requiere la identificación de individuos para estimar la densidad de
poblaciones de fauna silvestre y se basa en tres supuestos: a. El modelo se
construye con la tasa de encuentro de individuos asumiendo que los movimientos son
aleatorios e independientes unos de otros, b. las fotografías representan
contactos independientes entre individuos y cámaras, y c. la población es
cerrada (Rowcliffe et al. 2008). La población cerrada asume que, durante un muestreo breve, se
minimiza la rotación de la población objetivo por nacimientos, muertes,
inmigración y emigración (Karanth
et al. 2011; Mandujano Rodríguez 2011). La densidad poblacional (D) se calculó
como:
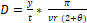
Donde, y = número de imágenes independientes,
t = esfuerzo de muestreo (trampas/noche), v = rango aproximado de
movimiento diario de la especie (Carbone
et al. 2005; Manzo et al. 2012), r = distancia de detección y ϴ = arco de
detección de la cámara (radianes) (Rowcliffe
et al. 2008).
Estimamos el parámetro (v) rango diario de movimiento del tapir andino mediante
el Modelo Alométrico (MA) propuesto por Carbone et al. (2005), el cual realiza una aproximación teórica entre el ámbito
hogareño y la disponibilidad de alimento. Este supuesto es usado para comparar
la relación entre los espacios bidimensionales y tridimensionales usados por
los mamíferos (Carbone
et al. 2005; Rovero y Marshall 2009). Sin embargo, el parámetro (v)
rango de movimiento diario así calculado presenta ciertas incertidumbres. Para
evaluar la sensibilidad de la densidad al parámetro de rango de movimiento
diario del tapir andino (v), se calculó la densidad utilizando tres valores de
(v) reportados en estudios previos, ordenados de menor a mayor: a. 0.964 km (Castellanos 2013), b. 7 km (estimado en este
estudio), y c. 14.6 km (Lizcano y Cavelier
2004b).
Para identificar la contribución de los
factores relacionados con la presencia del tapir andino se usaron, como ya
hemos indicado, variables de hábitat y de perturbación antrópica (Tabla 1).
Se construyeron modelos de ocupación agrupando los datos en cada uno de los dos
tipos de hábitats evaluados, bosque montano y páramo. Se estandarizaron todas
las variables continuas. Se generaron modelos candidatos para la ocupación (y) y la detectabilidad (p) de una temporada y una especie (Andrade-Ponce et al. 2021). Para ello se construyeron modelos para seleccionar la mejor
estructura de los efectos aleatorios mediante el estimador de Máxima
Verosimilitud Restringida (REML); posteriormente se generó un sub-set de
modelos posibles; finalmente se combinaron para generar una nueva selección de los
mejores modelos basado en el Criterio de Información de Akaike corregido para
muestra pequeñas (<2AICc) y Akaike peso (wi) (Andrade-Ponce
et al. 2021). Mediante
el paquete AICcmodavg (Mazerolle
2023), se evaluó el
ajuste (P) y el parámetro de sobredispersión (cˆ) con 1000
réplicas de arranque paramétrico. Cuando cˆ > 1, se usó cˆ
para inflar los errores estándar de los parámetros estimados (Burnham
y Anderson 2002; MacKenzie y Bailey 2004). Todos los análisis se realizaron con
el paquete Unmarked (Fiske y
Chandler 2011) en
RStudio versión 4.4.0 (R Core Team 2023).
Finalmente, se estimó el esfuerzo de
muestreo multiplicando el número total de trampas
cámara usadas por el total de días de muestreo encada sitio (Lira-Torres y Briones-Salas 2012). Este es entendido como un indicador de
la intensidad y el tiempo de la actividad de seguimiento llevada a cabo con
cámaras trampa en un área determinada (Lira-Torres y Briones-Salas 2012; Mandujano Rodríguez 2024).
Resultados
Densidad
En los 18 meses de seguimiento el
esfuerzo de muestreo alcanzó las 5192 trampas/noche (𝝁
64.9, SD±11.1) en bosque montano y 6131 trampas/noche (𝝁 76.6, SD±19.3) en páramo. Conseguimos 220 capturas independientes
de tapir andino, 179 en bosque montano y 41 en páramo. La densidad poblacional
estimada de tapir andino en bosque montano usando el MEA fue 0.24 individuos/km2;
(23.55 individuos/100km2), y en páramo fue 0.12 individuos/km2;
(11.59 individuaos/100km2) (Fig. 2). El análisis de
sensibilidad indicó que un aumento en el rango de movimiento diario del tapir
andino de v ≥ 7 km a v = 14
km no tuvo un efecto significativo en la estimación de la densidad. No
obstante, al utilizar un valor inferior a 7 km (v = 0.961 km), se observó un
incremento significativo en la densidad.
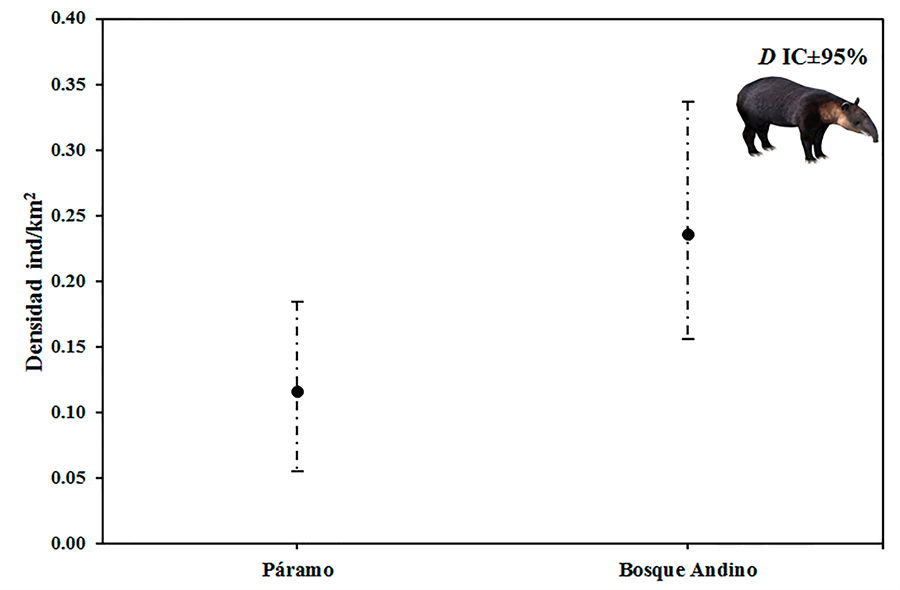
Figura 2. Densidad estimada para tapir andino
usando el parámetro v = rango aproximado de movimiento diario: v =7 km estimado
por este estudio; D = Densidad estimada; IC = Intervalo de confianza (líneas
punteadas).
Figure 2. Estimated density for Andean tapir using the parameter v =
approximate range of daily movement: v =7 km estimated by this study; D =
Estimated density; CI = Confidence Intervals (dotted lines).
Ocupación de hábitat
Las variables
predictoras seleccionadas para la ocupación variaron según el hábitat (Tabla 2). La ocupación promedio (yˆ) estimada para el tapir andino presentó
diferencias significativas entre hábitats, siendo mayor en el bosque montano (Tabla 3). En este hábitat la ocupación estuvo
positivamente asociada con la presencia de carnívoros, mientras que en el
páramo se relacionó con la cobertura vegetal (Tabla 3). Por otro lado, la detectabilidad promedio (pˆ)
estimada también presentó diferencias entre hábitats. En el bosque montano, se
relacionó de forma positiva con la cobertura vegetal, aunque fue inferior al
0.20 (Tabla 3). En el páramo, se mantuvo constante
y no estuvo influida por ninguna de las variables evaluadas (Tabla 3). El análisis de bondad de ajuste
indica que el modelo candidato con el menor <2AICc se ajusta bien
a los datos: bosque montano (P > 0.57, cˆ= 1), páramo (P =
0.4, cˆ= 1.09).
Tabla
2. Modelos
candidatos para tapir andino. Crv presencia de carnívoros; NDVI cobertura
vegetal; VIm distancia más cercana a caminos y vías; Evm elevación y Pam
distancia más cercana a zonas pobladas; (.) indica que no existe una variable
asociada para la Ψ o p.
Table 2. Candidate models for Andean tapir. Crv presence
of carnivores; NDVI vegetation cover; VIm closest distance to roads and tracks;
Evm elevation and Pam, closest distance to populated areas; (.) indicates that
there is no associated variable for Ψ or p.
Tabla 3. Coeficientes β estimado para el mejor modelo de ocupación y detectabilidad en
cada hábitat. Ocupación (ψ^) y detectabilidad (p^)
media estimada. Crv, presencia de carnívoros; NDVI
cobertura vegetal.
Table
3. β coefficients for the best model for occupancy
and detectability in each habitat. Estimated mean occupancy (ψ^) and
detectability (p^). Crv abundance of carnivores; NDVI vegetation cover.
Discusión
Densidad
Nuestra aproximación con fototrampeo ha
demostrado ser efectiva para detectar al tapir andino, una especie rara y
esquiva (Cruz et al.
2014; Zapata-Ríos y Branch 2016; Schank et al. 2017, 2019; Nakashima
et al. 2018).
Nuestras estimaciones con el Modelo de Encuentros Aleatorios proporcionó
resultados similares a los obtenidos con técnicas como la telemetría, que
logísticamente es más complicada y costosa de implementar, y con técnicas
indirectas (Acosta
et al. 1996; Downer 1996; Lizcano y Cavelier 2000b; Castellanos
2013) (Tabla A1 del Anexo). El MEA permitió estimar la densidad poblacional del tapir
andino sin la necesidad de identificar individuos algo especialmente complicado
para esta especie (Rowcliffe
et al. 2008; Rovero y Marshall 2009; Manzo et al.
2012; Carbajal-Borges et al. 2014).
Pese a que la densidad fue mayor en
bosque montano, el cual es considerado un hábitat crítico y con mayor
abundancia de tapir andino, por ofrecer mayor cantidad de alimento y refugio
que el páramo (Downer 2001; Bermúdez
y Reyes Puig 2011), la densidad estimada para tapir andino no mostró
diferencias estadísticamente significativas entre estos hábitats (Fig. 2).
Consideramos que este resultado puede ser consecuencia de las migraciones
altitudinales anuales del tapir andino entre bosque montano y páramo causadas
por la estacionalidad climática (Acosta
et al. 1996; Downer 1996). En
estudios previos realizados en hábitats similares en Ecuador (Downer 1996; Castellanos
2013) y Colombia (Acosta
et al. 1996; Lizcano y Cavelier 2000b) se encontraron densidades de tapir
andino similares a las reportados aquí (Tabla A1 del Anexo).
En un
estudio realizado en el páramo de Papallacta, Ecuador, se detectaron siete
individuos en un área de 1442 ha, lo que supone una densidad de 0.29
individuos/km2 (Castellanos 2013) que es algo mayor a la
reportada en este estudio y en otros (Tabla A1 del Anexo). Aunque las técnicas de muestreo son diferentes,
creemos que las diferencias en la densidad se deben a las características de
las localidades evaluadas. Las densidades estimadas por Downer
(1996) y Acosta et
al. (1996) no
consideran diferencias entre hábitats y consideran el páramo y el bosque
montano como una sola área de muestreo. Sin embargo, la densidad de tapir
andino obtenida por Lizcano y Cavelier (2000a) solo en bosque montano son similares
a las registradas por Downer (1996) y Acosta et al. (1996) (Tabla A1 del Anexo). Estos resultados sugieren una mayor densidad del
tapir andino en los bosques montanos (Downer 1996; Padilla et al. 2010), lo cual estaría relacionado con la mayor
productividad del bosque, con la oferta de recursos alimenticios y con la protección
contra depredadores (Lizcano
y Cavelier 2000b; Downer 2001; Bermúdez
y Reyes Puig 2011).
Si bien el MEA es efectivo para estimar la densidad del tapir andino (Manzo et al.
2012), persiste la incertidumbre asociada al rango de
movimiento diario de la especie (v; km) (Carbajal-Borges
et al. 2014).
Esta variación en la densidad del tapir andino como efecto del parámetro (v; km) se muestran en este estudio (Fig. A1 del Anexo). Un efecto similar fue reportado en el caso del tapir
mesoamericano Tapirus bairdii (Carbajal-Borges
et al. 2014). Esta
incertidumbre es esperable, dado que un rango de movimiento diario reducido
incrementa la probabilidad de detección de la especie en las cámaras trampa. Sin
embargo, el rango de movimiento diario real del tapir andino sigue siendo
difícil de determinar con precisión.
Uno de los valores usados para el
análisis de sensibilidad de 14.4 km/día (Lizcano
y Cavelier 2004b), no
es representativo de la población de tapir andino, dado que es un resultado que
corresponde a un solo individuo en seis meses de muestreo. Sabemos que este
parámetro varía con el tipo de hábitat, género, edad del individuo y
estacionalidad (Lizcano
y Cavelier 2004b; Castellanos 2013). Otros autores obtuvieron estimaciones
más robustas del rango de movimiento diario para otras especies no relacionadas
con el tapir andino, basadas en radio-tracking y modelo alométrico, con
un número mayor de individuos muestreados 4 a 14, respectivamente (Rovero y
Marshall 2009; Manzo et al. 2012). Obtener mejores estimaciones sobre el rango de movimiento diario
para el tapir andino permitirá despejar esta duda y mejorar la estimación de su
densidad.
Variables que afectan la
presencia del tapir andino
Nuestros modelos de ocupación muestran
que los factores determinantes varían entre hábitats. En el bosque montano, la
presencia de carnívoros y la cobertura vegetal tienen una relación positiva con
su ocupación y detectabilidad. Mientras que, en el páramo, la ocupación está
relacionada positivamente con la cobertura vegetal, y, la detectabilidad no se
relacionó con ninguna variable evaluada (Fig. 3).
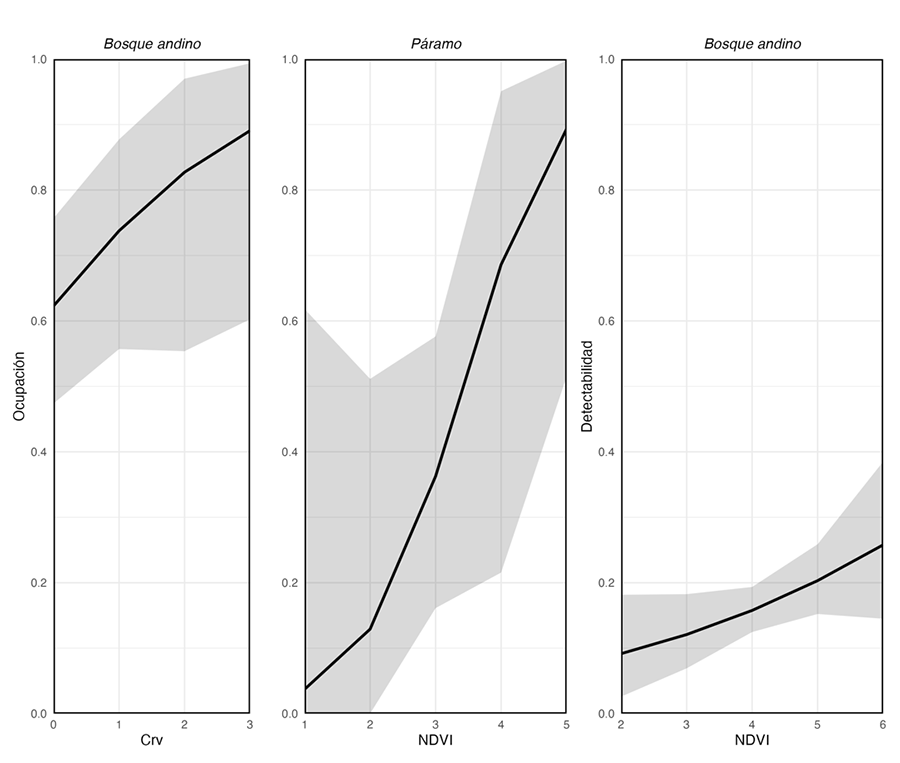
Figura 3. Ocupación y detectabilidad del tapir andino en bosque montano y
páramo estimados con nuestros modelos. Crv, presencia
de carnívoros; NDVI, cobertura vegetal.
Figure 3. Occupation and predictive detectability of Andean tapir in montane
forest and páramo. Crv, presence of carnivores; NDVI, vegetation cover.
A pesar de que la relación positiva entre
la ocupación del tapir andino y la presencia de carnívoros en el bosque montano
parece contradictoria, los registros de depredación del tapir andino son
escasos. Peyton (1980) documentó la depredación de
un tapir andino por oso andino en Perú, mientras que Downer (1996) informó de la presencia de pelos de tapir andino en las heces de
oso andino y puma. Rodríguez
y colaboradores (2014) registraron en una cámara
trampa el ataque de un oso andino a un tapir andino adulto. En Colombia se
detectaron cicatrices de garras en individuos de tapir andino (Pisso-Florez
et al. 2021) . Los bosques montanos son altamente productivos en recursos
(alimento, refugio, saladeros y agua), tanto para el tapir andino (Lizcano
y Cavelier 2000b, 2004a; Downer 2001) como para otras especies de grandes
mamíferos (Zapata-Ríos
y Branch 2018). Esto explicaría el efecto positivo de la presencia de
carnívoros, los cuales se alimentan de otros herbívoros, y no solo del tapir
andino.
Esta mayor
productividad del bosque se refleja en la variable de cobertura vegetal como un
predictor positivo de la detectabilidad en el bosque y en la ocupación del
tapir andino en el páramo (Fig.
3). Esta relación se debe probablemente a la elevada demanda
energética del tapir andino (Bermúdez
y Reyes Puig 2011) y a la
disponibilidad de especies vegetales de las que se alimenta (Acosta
et al. 1996). En el páramo, la relación con la cobertura
vegetal está sujeta a la existencia de parches de bosque, ya que estos ofrecen al tapir andino sitios seguros de protección y refugio (Acosta
et al. 1996; Downer 1996; Padilla et al. 2010). Nuestros resultados son consistentes
con los de Downer (1996), quien registró un uso promedio del tapir andino en paramo superior
al 6% frente al 29% del bosque montano. Sin embargo, en Ecuador, se reportó el
uso exclusivo del páramo herbáceo por el tapir andino (Castellanos
2013), pese a las
limitaciones de productividad y refugio de este hábitat .
Estudios realizados en Ecuador han
demostrado que la presencia de perros ferales altera el patrón de actividad y
reduce la abundancia del tapir andino (Tapirus pinchaque) (Zapata-Ríos y Branch 2016). Castellanos et al. (2022)
documentaron un caso de ataque de perros ferales, con transmisión de
enfermedades como la rabia, que resultó en la muerte de un individuo. En
Colombia, se ha reportado el ataque de perros domésticos a un tapir andino (Cepeda-Duque et al. 2024). La presencia de perros
ferales y domésticos en el área de distribución de esta especie representa una
amenaza creciente, con impactos potenciales en su ecología y viabilidad
poblacional, particularmente en zonas cercanas a centros poblados. No obstante,
en este estudio no se detectó la presencia de estos animales. En Perú, la ocupación del tapir andino estuvo afectada de forma positiva
con la distancia a carreteras y negativamente con la topografía (Mena et al.
2020). En este estudio no se detectó una condición similar,
probablemente por el nivel de aislamiento y buen estado de conservación de las
áreas evaluadas. Es razonable esperar que la ocupación del tapir andino
sea mayor mientras incrementa la distancia a vías y poblados, condición derivada de ser una especie tímida, solitaria y que
frecuenta zonas de bosque y páramos en buen estado de conservación (Padilla
et al. 2010).
Implicaciones para la conservación
La densidad estimada en este estudio y
por otros autores sugieren que esta especie es naturalmente rara, con
densidades poblacionales bajas. En comparación con la densidad estimada para
el tapir amazónico Tapirus terrestris de 0.8 individuo/km2 (Ferreguetti
et al. 2017) y tapir
mesoamericano con 0.39 individuo/km2 (Botello et al. 2017). Los resultados, también
muestran que las variables explicativas como la presencia de carnívoros y la
cobertura vegetal son importantes e influyen positivamente en la ocupación y
detectabilidad del tapir andino respectivamente. Esto muestra la importancia
de las áreas protegidas como refugios para el tapir andino y la fauna silvestre
(Ortega-Andrade
et al. 2015; Mestanza-Ramón et al. 2021). También pone de manifiesto la
susceptibilidad de la especie a cambios drásticos en el uso del suelo, la
deforestación, el avance de la frontera agrícola y la presencia de especies
invasoras, como perros ferales (Ortega-Andrade
et al. 2015; Zapata-Ríos y Branch 2016, 2018). La
incidencia de factores que afectan la ocupación y detectabilidad del tapir
andino que varían espacial y temporalmente subraya la necesidad de estrategias
de conservación adaptativas a las distintas realidades.
Conclusiones
Este estudio contribuye a la comprensión
de la ecología y demografía del tapir andino en los Andes del norte de Ecuador
en sus dos principales hábitats. Si bien no se observaron diferencias
significativas entre hábitats, nuestros datos subrayan la baja densidad del
tapir andino y su dependencia de áreas bien conservadas. En términos de
ocupación, la presencia de carnívoros afecta positivamente en el bosque
montano, mientras que la cobertura vegetal lo hace en el páramo. Las densidades
de tapir andino indican el valor de las áreas protegidas como refugios críticos
para el tapir andino y la fauna silvestre asociada. Finalmente se señalan los
aspectos que requieren ser abordados para afinar, corregir y garantizar la
efectividad de las medidas de manejo y conservación: a) evaluar la
incertidumbre del rango diario de movimiento del tapir andino; b) evaluar las
variaciones demográficas en el tiempo; c) involucrar a las comunidades dentro
de su área de distribución en monitoreo del tapir andino; d) campañas de manejo
y esterilización de perros domésticos en centros poblados y e) remoción de
perros sin dueños en las áreas de distribución del tapir andino.
Contribución de los autores
Conceptualización: Carlos
Urgilés-Verdugo; Metodología: Carlos
Urgilés-Verdugo; Investigación: Carlos
Urgilés-Verdugo, Freddy Gallo-Viracocha; Análisis formal: Carlos Urgilés-Verdugo, Michael S. Esbach; Redacción-borrador
inicial: Carlos Urgilés-Verdugo; Curación de datos: Carlos Urgilés-Verdugo, Michael S. Esbach; Redacción-Revisión
y edición: Carlos Urgilés-Verdugo, Freddy Gallo-Viracocha,
Michael S. Esbach, Adrián Escudero; Obtención de fondos: Carlos
Urgilés-Verdugo; Administración del proyecto: Carlos Urgilés-Verdugo; Supervisión:
Adrián Escudero, Michael S. Esbach.
Disponibilidad de datos
Los datos de este estudio están
disponibles en el repositorio digital Zenodo (https://doi.org/10.5281/zenodo.13760478).
Financiación, permisos requeridos, potenciales
conflictos de interés y agradecimientos
Este proyecto de investigación se realizó
gracias al apoyo de EcoFondo-Ecuador. Agradecemos al Ministerio del Ambiente
del Ecuador (MAE) por la extensión de los permisos de investigación
(047-10-IC-FAU / FLO-DPN / MA y 014-IC-FAU / -DPS / MA); a Leopoldo Gómez,
Abraham Romero y Ricardo Urcuango, guardaparques del MAE, por el apoyo
proporcionado. También agradecemos a Jorge y Arturo Benavidez y Leopoldo Gómez
por su apoyo en la comunidad de Cuyuja; a la Fundación para la Supervivencia
Cofán (CSF) y sus guardaparques por su apoyo en la comunidad de Monte Olivo; a
Norberto Parión y Fabian Ascanta por su apoyo en la comunidad de Oyacachi. Los
comentarios de Galo Zapata-Ríos, Diego Lizcano y Carlos Iván Espinosa mejoraron
las revisiones iniciales de este manuscrito.
El autor y los coautores declaran que no
tienen ningún conflicto de intereses.
Referencias
Acosta, H., Cavelier, J., Londoño, S. 1996. Aportes
al Conocimiento de la Biología de la Danta de Montaña, Tapirus pinchaque, en
los Andes Centrales de Colombia. Biotropica 28: 258-266. https://doi.org/10.2307/2389080
Andrade-Ponce, G.,
Cepeda-Duque, J.C., Mandujano, S., Velásquez-C, K.L., Gómez-Valencia, B.,
Lizcano, D.J. 2021. Modelos de ocupación para datos de cámaras trampa: de los
conceptos a la interpretación. Mammalogy Notes 7: 1-23. https://doi.org/10.47603/mano.v7n1.200
Baquero, F., Sierra, R.,
Ordóñez, L., Tipán, M., Espinosa, L., Rivera, M.B., Soria, P. 2004. La
vegetación de los Andes del Ecuador. Memoria explicativa de los mapas de
vegetación: potencial y remanente a escala 1:250.000 y del modelamiento
predictivo con especies indicadoras. EcoCiencia/CESLA/Cor- poración
EcoPar/MAG SIGAGRO/CDC - Jatun Sacha/División Geográfica - IGM, Quito.
Bermúdez, D.K., Reyes
Puig, J.P. 2011. Dieta del tapir de montaña (Tapirus pinchaque) en
tres localidades del corredor ecológico Llangantes – Sangay. Boletín
Técnico, Serie Zoológica 10 (7): 1-13. https://journal.espe.edu.ec/ojs/index.php/revista-serie-zoologica/article/view/1444
Botello, F.,
Romero-Calderón, A.G., Sánchez-Hernández, J., Hernández, O., López-Villegas,
G., Sánchez-Cordero, V. 2017. Densidad poblacional del tapir centroamericano (Tapirella
bairdii) en bosque mesófilo de montaña en Totontepec Villa de Morelos,
Oaxaca, México. Revista Mexicana de Biodiversidad 88: 918-923. https://doi.org/10.1016/j.rmb.2017.10.016
Burnham, K.P., Anderson,
D.R. 2002. Model Selection and Inference: A Practical
Information-Theoretic Approach. 2nd ed.
Springer-Verlag, New York.
Carbajal-Borges, J.P., Godínez-Gómez, O., Mendoza, E. 2014. Density, Abundance and
Activity Patterns of the Endangered Tapirus Bairdii in One of its Last
Strongholds in Southern Mexico. Tropical Conservation Science 7:
100-114. https://doi.org/10.1177/194008291400700102
Carbone, C., Cowlishaw, G., Isaac, N.J.B., Rowcliffe, J.M. 2005. How Far Do
Animals Go? Determinants of Day Range in Mammals. The American Naturalist
165: 290-297. https://doi.org/10.1086/426790
Castellanos, A. 2013. Iridium/GPS Telemetry to Study Home Range and Population
Density of Mountain Tapirs in the Rio Papallacta Watershed, Ecuador. Tapir Conservation (The Newsletter of
the IUCN/SSC Tapir Specialist Group) 22 (31): 20-25. https://iucntsg.wpengine.com/wp-content/uploads/2017/03/Tapir-Conservation-News-Dec-2013.pdf
Castellanos, A.X.,
Brito, J., Castellanos, F. 2020. Registros de Distribución de Tapir Andino, Tapirus
pinchaque en el occidente de Ecuador. Boletín Técnico, Serie Zoológica
15 (16): 1-4. https://journal.espe.edu.ec/ojs/index.php/revista-serie-zoologica/article/view/1629
Castellanos, A., Dadone, L., Ascanta, M., Pukazhenthi, B. 2022. Andean tapir (Tapirus pinchaque) social groups and calf
dispersal patterns in Ecuador. Boletín Técnico,
Serie Zoológica 17: 09-14. https://journal.espe.edu.ec/ojs/index.php/revista-serie-zoologica/article/view/2858
Cepeda-Duque JC,
Arango-Correa E, Frimodt-Møller C, Lizcano DJ. 2024. Howling
shadows: First report of domestic dog attacks on globally threatened mountain tapirs in high Andean cloud
forests of Colombia. Neotropical Biology and Conservation 19:25-33.
https://doi.org/10.3897/neotropical.19.e117437
Cruz, P., Paviolo, A., Bó, R.F., Thompson, J.J., Di Bitetti, M.S. 2014. Daily activity patterns and habitat use of the lowland tapir
(Tapirus terrestris) in the Atlantic Forest. Mammalian Biology 79:
376-383. https://doi.org/10.1016/j.mambio.2014.06.003
Damuth, J. 1981. Population sensity and body size in animmals. Nature
290: 699-700. https://doi.org/10.1038/290699a0
De la Torre, L., Muriel,
P., Balslev, H. 2006. Etnobotánica en los Andes del Ecuador. En: Moraes, M.,
Øllgaard, B., Kvist, L.P., Borchsenius, F., Balslev, H. (eds.), Botánica
Económica de los Andes Centrales, pp. 246-267. Universidad Mayor de San
Andrés, La Paz.
Downer, C.C. 1996. The mountain tapir, endangered ‘flagship’ species
of the high Andes. Oryx 30: 45-58. https://doi.org/10.1017/S0030605300021384
Downer, C.C. 2001. Observations on the diet and habitat of the mountain
tapir (Tapirus pinchaque). Journal of Zoology 254: 279-291. https://doi.org/10.1017/S0952836901000796
Efford, M. 2004. Density estimation in live‐trapping
studies. Oikos 106: 598-610. https://doi.org/10.1111/j.0030-1299.2004.13043.x
Ferreguetti, Á.C., Tomás, W.M., Bergallo, H.G. 2017. Density, occupancy, and
detectability of lowland tapirs, Tapirus terrestris, in Vale Natural Reserve,
southeastern Brazil. Journal of Mammalogy 98: 114-123. https://doi.org/10.1093/jmammal/gyw118
Fiske,
I.J., Chandler, R.B. 2011. unmarked: An R Package
for Fitting Hierarchical Models of Wildlife Occurrence and Abundance. Journal
of Statistical Software 43: 1-23. https://doi.org/10.18637/jss.v043.i10
Hofstede, R., Coppus, R., Mena Vásconez, P., Segarra, P., Wolf, J., Sevink,
J. 2002. El estado de conservación de los páramos de pajonal en el
Ecuador. ECOTROPICOS (Sociedad Venezolana de Ecología) 15(1):
3-18. https://www.researchgate.net/publication/260423686
Jiménez, C.F., Quintana, H.,
Pacheco, V., Melton, D., Torrealva, J., Tello, G. 2010. Camera
trap survey of medium and large mammals in a montane rainforest of northern
Peru. Revista Peruana de Biología 17: 191-196. https://doi.org/10.15381/rpb.v17i2.27
Karanth, K.U., Ullas
Karanth, K., Nichols, J.D., Samba Kumar, N. 2011. Estimating
Tiger Abundance from Camera Trap Data: Field Surveys and Analytical Issues. In:
O’Connell, A. F., Nichols, J.D., Karanth, K.U. (eds.), Camera Traps in
Animal Ecology, pp. 97-117. Springer Japan, Tokyo. https://doi.org/10.1007/978-4-431-99495-4_7
Lira-Torres, I.,
Briones-Salas, M. 2012. Abundancia relativa y patrones de actividad de los
mamíferos de los Chimalapas, Oaxaca, México. Acta Zoológica Mexicana (N.S.)
28: 566-585. https://doi.org/10.21829/azm.2012.283859
Lizcano, D.J., Cavelier,
J. 2000a. Daily and seasonal activity of the mountain
tapir (Tapirus pinchaque) in the Central Andes of Colombia. Zoological
Society of London 429-435. https://doi.org/10.1017/S0952836900000182
Lizcano, D.J., Cavelier, J. 2000b. Densidad Poblacional y
Disponibilidad de Hábitat de la Danta de Montaña (Tapirus pinchaque) en
los Andes Centrales de Colombia1. Biotropica 32: 165-173. https://doi.org/10.1111/j.1744-7429.2000.tb00458.x
Lizcano, D.,
Cavelier, J. 2004a. Características Químicas de salados y hábitos alimenticios
de la Danta de montaña (Tapirus pinchaque Roulin, 1829) en los Andes Centrales
de Colombia. Mastozoología Neotropical 11: 193-201.
Lizcano, D.J., Cavelier, J. 2004b. Using GPS Collars to Study Mountain
Tapirs (Tapirus pinchaque) in the Central Andes of Colombia. Tapir
Conservation (The Newsletter of the IUCN/SSC Tapir Specialist Group)
13 (2): 18-23. https://iucntsg.wpengine.com/wp-content/uploads/2017/03/Tapir-Conservation-News-Dec-2004.pdf
Lizcano, D.J., Pizarro, V.,
Cavelier, J., Carmona, J. 2002. Geographic distribution and
population size of the mountain tapir (Tapirus pinchaque) in Colombia. Journal
of Biogeography 29: 7-15. https://doi.org/10.1046/j.1365-2699.2002.00654.x
MacKenzie,
D.I., Bailey, L.L. 2004. Assessing the fit of
site-occupancy models. Journal of Agricultural, Biological, and
Environmental Statistics 9: 300-318. https://doi.org/10.1198/108571104X3361
Mackenzie,
D.I., Royle, J.A. 2005. Designing occupancy
studies: general advice and allocating survey effort. Journal of
Applied Ecology 42: 1105-1114. https://doi.org/10.1111/j.1365-2664.2005.01098.x
Mandujano Rodríguez,
S. 2011. Ecología de poblaciones aplicada al manejo de fauna silvestre:
cuatro conceptos (N, ʎ, MSY, Pe). Instituto Literario de Veracruz S. C.,
Veracruz, México.
Mandujano Rodríguez,
S. 2024. Índice de abundancia relativa y tasa de encuentro con trampas cámara. Mammalogy Notes 10: 389. https://doi.org/10.47603/mano.v10n1.389
Manzo, E., Bartolommei, P., Rowcliffe, J.M., Cozzolino, R. 2012.
Estimation of population density of European pine marten in central Italy using
camera trapping. Acta Theriologica 57: 165-172. https://doi.org/10.1007/s13364-011-0055-8
Mazerolle, M.J. 2023. AICcmodavg: Model Selection and Multimodel Inference
Based on (Q)AIC(c). 2.3-3. https://doi.org/10.32614/CRAN.package.AICcmodavg
Mena, J.L., Yagui, H., La Rosa,
F., Pastor, P., Rivero, J., Appleton, R. 2020. Topography and
disturbance explain mountain tapir (Tapirus pinchaque) occupancy at its
southernmost global range. Mammalian Biology 100: 231-239. https://doi.org/10.1007/s42991-020-00027-9
Mena Vásconez, P.,
Hofstede, R. 2006. Los páramos ecuatorianos. En: Moraes, M., Øllgaard, B.,
Kvist, L.P., Borchsenius, F., Balslev, H. (eds.), Botánica Económica de los
Andes Centrales, pp. 91-109. Universidad Mayor de San Andrés, La Paz,
Bolivia.
Mestanza-Ramón, C.,
Herrera Feijoo, R.J., Chicaiza-Ortiz, C., Gaibor, I.D., Mateo, R.G. 2021. Estimation of Current and Future Suitable Areas for Tapirus
pinchaque in Ecuador. Sustainability 13: 11486. https://doi.org/10.3390/su132011486
Ministerio del
Ambiente del Ecuador. 2013. Sistema de Clasificación de los Ecosistemas
del Ecuador Continental. Subsecretaría de Patrimonio Natural, Quito,
Ecuador.
Nakashima, Y., Fukasawa,
K., Samejima, H. 2018. Estimating animal density without
individual recognition using information derivable exclusively from camera
traps Stephens, P. (ed.). Journal of Applied Ecology 55: 735-744. https://doi.org/10.1111/1365-2664.13059
O’Brien, T.G., Kinnaird, M.F., Wibisono, H.T. 2003. Crouching tigers,
hidden prey: Sumatran tiger and prey populations in a tropical forest
landscape. Animal Conservation 6: 131-139. https://doi.org/10.1017/S1367943003003172
O’Connell, A.F., Nichols, J.D., Ullas Karanth, K. (eds.). 2011. Camera
Traps in Animal Ecology. Springer Japan, Tokyo, Japan. https://doi.org/10.1007/978-4-431-99495-4
Ortega-Andrade, H.M.,
Prieto-Torres, D.A., Gómez-Lora, I., Lizcano, D.J. 2015. Ecological
and Geographical Analysis of the Distribution of the Mountain Tapir (Tapirus
pinchaque) in Ecuador: Importance of protected areas in future scenarios of
global warming. PLOS ONE 10: 1-20. https://doi.org/10.1371/journal.pone.0121137
Padilla, M., Dowler, R.C., Downer, C.C. 2010. Tapirus pinchaque
(Perissodactyla: Tapiridae). Mammalian Species 42: 166-182. https://doi.org/10.1644/863.1
Peyton, B. 1980. Distribution, and Food Habits of Spectacled Bears, Tremarctos
ornatus, in Peru. Journal of Mammalogy 61: 639-652. https://doi.org/10.2307/1380309
Pisso-Florez, G.A., Gómez-Lora, I., Vela-Vargas, I.M., Pizo, H., Bedoya Dorado,
I., Ramírez-Chaves, H.E. 2021. What’s on the menu? A presumed attack of Andean
bear on a Mountain tapir at the Puracé National Natural Park, Colombia. Neotropical
Biology and Conservation 16: 19-25. https://doi.org/10.3897/neotropical.16.e57140
Quichimbo, P., Tenorio,
G., Borja, P., Cárdenas, I., Crespo, P. 2012. Efectos sobre las propiedades
físicas y químicas de los suelos por el cambio de la covertura vegetal y udo
del suelo: páramo de Quinsacocha al sur del Ecuador. Sociedad Colombiana de
la Ciencia del Suelo 42: 138-153.
R Core Team. 2023. R: A language and environment for statistical computing (version 4.3.0). R Foundation for Statistical Computing, Vienna,
Austria. https://www.r-project.org/
Rodriguez, A., Gomez, R., Moreno, A., Cuellar, C., Lizcano, D.J. 2014. Record
of a mountain tapir attacked by an Andean bear on a camera trap. Tapir
Conservation (The Newsletter of the IUCN/SSC Tapir Specialist Group)
23: 25-26. https://iucntsg.wpengine.com/wp-content/uploads/2017/03/Tapir-Conservation-News-July-2014.pdf
Rovero,
F., Marshall, A.R. 2009. Camera trapping
photographic rate as an index of density in forest ungulates. Journal of
Applied Ecology 46: 1011-1017. https://doi.org/10.1111/j.1365-2664.2009.01705.x
Rowcliffe, J.M., Field, J., Turvey, S.T., Carbone, C. 2008. Estimating animal
density using camera traps without the need for individual recognition. Journal
of Applied Ecology 45: 1228-1236. https://doi.org/10.1111/j.1365-2664.2008.01473.x
Schank, C.J., Cove, M.V., Kelly, M.J., Mendoza, E., O’Farrill, G., Reyna‐Hurtado,
R., Meyer, N., et al. 2017. Using a novel model approach to assess the
distribution and conservation status of the endangered Baird’s tapir Thuille,
W. (ed.). Diversity and Distributions 23: 1459-1471. https://doi.org/10.1111/ddi.12631
Schank, C.J., Cove, M.V., Kelly, M.J., Nielsen, C.K., O’Farrill, G.,
Meyer, N., Jordan, C.A., et al. 2019. A Sensitivity Analysis of the
Application of Integrated Species Distribution Models to Mobile Species: A Case
Study with the Endangered Baird’s Tapir. Environmental Conservation
46: 184-192. https://doi.org/10.1017/S0376892919000055
Tejedor Garavito,
N., Álvarez, E., Arango Caro, S., Araujo Murakami, A., Blundo, C., Boza
Espinoza, T.E., La Torre Cuadros, M.A., et al. 2012. Evaluación del estado
de conservación de los bosques montanos en los Andes tropicales. Ecosistemas
21(1-2): 148-166. Recuperado a partir de https://www.revistaecosistemas.net/index.php/ecosistemas/article/view/34
Tirira, D., Urgilés-Verdugo,
C., Tapia, A., Cajas-Bermeo, C.A., Izurieta V, X., Zapata-Ríos, G. 2019. Tropical Ungulates of Ecuador: An Update of the State of Knowledge.
In: Gallina-Tessaro, S. (ed.), Ecology and Conservation of Tropical
Ungulates in Latin America, pp. 217-271. Springer Nature,
Switzerland. https://doi.org/10.1007/978-3-030-28868-6_11
Urgilés-Verdugo, C.,
Cisneros-Vidal, R., Molina Proaño, S., Ordoñez-Delgado, L., Reyes-Puig, J.,
Sánchez-Karste, F., Tapia, A., et al. 2021. Tapir andino (Tapirus
pinchaque). En: Tirira, D. (ed.), Libro Rojo de mamíferos del Ecuador,
(3ª ed.). Asociación Ecuatoriana de Mastozoología, Fundación Mamíferos y
Conservación, Pontificia Universidad Católica del Ecuador y Ministerio del
Ambiente, Agua y Transición Ecológica del Ecuador, Quito, Ecuador. https://aem.mamiferosdelecuador.com/images/pdf/Tirira_2021-Lista_roja_de_los_mamiferos_del_Ecuador_2021.pdf
Zapata-Ríos, G., Branch,
L.C. 2016. Altered activity patterns and reduced abundance
of native mammals in sites with feral dogs in the high Andes. Biological
Conservation 193: 9-16. https://doi.org/10.1016/j.biocon.2015.10.016
Zapata-Ríos, G., Branch, L.C. 2018. Mammalian carnivore
occupancy is inversely related to presence of domestic dogs in the high Andes
of Ecuador. PLOS ONE 13: 1-17. https://doi.org/10.1371/journal.pone.0192346
Anexo / Appendix
Tabla A1. Comparación
de la densidad estimada para tapir andino entre estudios realizados, hábitats y
metodologías.
Table A1. Comparison of estimated density for Andean tapir among studies,
habitats, and methodologies.
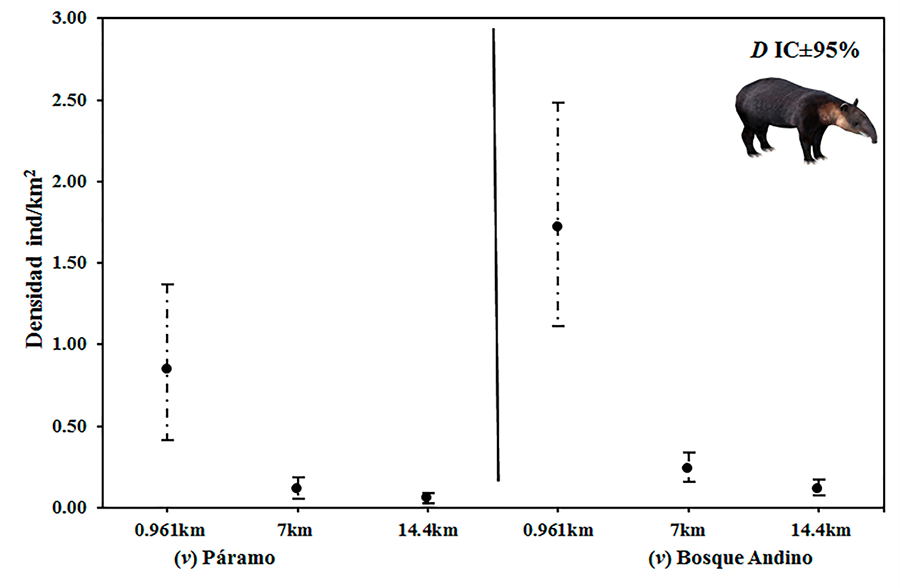
Figura A1. Análisis de sensibilidad de la
densidad estimada para tapir andino con base en el parámetro v = rango de
movimiento diario: v =7 km (éste estudio); v =14.4 km (Lizcano y Cavelier 2000b) v = 0.961 km (Castellanos 2013); D = Densidad estimada; IC = Intervalos de
Confianza al 95% (líneas punteadas).
Figure
A1. Sensitivity
analysis of the estimated density for Andean tapir based on the parameter v =
daily movement range: v =7 km (this study); v =14.4 km (Lizcano
and Cavelier 2000b) v =
0.961 km (Castellanos 2013);
D = Estimated density; CI = 95% Confidence Intervals (dotted lines).